Her er en typisk darwinistisk påstand: "Det som evolusjon har, er hva hvilken som helst god vitenskapelig påstand har: bevis og mye av det. Evolusjon er støttet av et bredt spekter av observasjoner, gjennom feltene genetikk, anatomi, økologi, dyreatferd, paleontologi og andre. Om du ønsker å utfordre evolusjonsteorien, må du adressere de bevis som finnes. Du må vise at bevis for den enten er feilaktig eller irrelevant, eller passer bedre til en annen teori. For å gjøre dette, må en kjenne både teorien og bevisene." (talkorigins.org/faqs/faq-misconceptions.html)
I dette avsnittet undersøkes bevis for felles avstamning, allment kjent som evolusjon. Den moderne profesjonelle litteraturen synes ikke å å bry seg mye om bevis. Kanskje er det fordi de betrakter sannhetsspørsmålet å være avgjort, noe som gjør en systematisk diskusjon av det overflødig. Gjennomlesning av litteraturen viser eks. på nye oppdagelser som sies å være bevis for felles avstamning, men eks. på generell drøfting av bevisene er fåtallige. Systematisk diskusjon av bevis finnes flest i bøker skrevet for å overbevis legfolk om sannheten av evolusjon. En av de bedre av disse, er en bok utkommet i 2008, en publikasjon fra National Academy of Science (NAS) og tre senere bøker (Coyne 2009, Dawkins 2009, Rogers 2011).
Bilde 1. Eks. på sirkelslutning om homologi
Bevisene inkluderer fossiler, fylogenetiske (slektskaps) trær, biogeografiske data, rudimentære organer og eks. på evolusjon under utvikling. Ut fra det som tidligereergåttgjennom i denne boka, vil jeg vise hvorfor disse argumentene for felles avstamning er feilaktige. Jeg vil også presentere en alternativ forklaring for bevisene, som jeg tror er bedre enn den om felles avstamning. Det vil vise seg at det ikke er noen gode bevis for felles avstamning.
Som nevnt tidligere, er et avgjørende kjennetegn ved livet, informasjonen det innebærer og det evne til å benytte informasjonen til å fungere og reprodusere. Hovedutfordringen til felles avstamning er å gjøre rede for hvordan den informasjonen er blitt bygd opp gjennom en naturlig prosess. Når alt kommer til alt, så inneholder nåtidige organismer som fisker, elefanter og mennesker åpenbart mye mer informasjon enn det som kunne ha vært i den formodentlig primitive cellen, som alt liv hevdes å ha utviklet seg fra. Det har ikke vært vist hvordan naturlige prosesser kunne bygge denne informasjonen, noe som måtte gjøres om vi forventes å betrakte felles avstamning som en skikkelig vitenskapelig teori.
Kan neo-darwinistisk teori gjøre rede for oppbygningen av informasjon? Har darwinister noensinne vist hvordan informasjonen i levende organismer kan ha blitt bygd opp, ved mekanismene til tilfeldige mutasjoner og naturlig seleksjon? Bidrar noe av det vanlige nevnte 'bevis' til oppbygning av informasjon? Svaret til disse tre spørsmålene er : Nei.
Det fylogenetiske tre som bevis for felles avstamning
Om 'felles-avstamnings' paradigmet var sant, burde en kunne være i stand til å konstruere et tre med organismer, som viser avstamningsmønsteret. Et fylogenetisk tre, som av og til kalles livets tre, er et diagram som viser hva som menes å være evolusjonær nedstigning av dyr og planter. Et tre kan konstrueres ved å følge ett spesielt sett organer gjennom hvilket en kan følge ett spesielt sett organer eller ulike slags dyr (eller planter). En kunne f.eks. gruppere dyr etter likheten i lemmene deres, eller fordøyelsessystemet, eller blod-sirkulasjons-systemet. Om en brukte lemmer som et karakteristika, ville grupperingen ta form av et tre hvor dyr med mest like lemmer ble plassert på samme tre-grein, mens de med mindre like lemmer ville plasseres på ulike greiner. Måten grenene forgrener seg, er ment å vise hvordan dyr utviklet seg fra hverandre. Alternativt kunne en som nevnt gruppere etter f.eks. blod-sirkulasjons-system. Om trær som konstrueres ved å sammenlikne atskillig ulike organer eller fysiologiskes systemer er identiske, så kunne en ha noe tillit til at treet virket meningsfullt. Om en ved å sammenlikne alle mulige biologiske trekk, ender opp med samme tre, så kunne treet representere en objektiv virkelighet. R. Dawkins (2009, s.321 ff.) tilbød hva han kalte et 'mektig bevis' for felles avstamning, basert på den (formodede) eksistens av et konsistent fylogenetisk tre.
I følge neo-darwinistisk paradigme, skulle dyr som har lignende karakteristika, være på samme grein i treet. Alle dyr som deler ett bestemt karakteristika skulle også dele en seneste felles stamfar, som var den første til å ha denne karakteristikken. Dawkins mener at evnen til å arrangere alle levende organismer i ett felles tre, ville være et bevis på felles avstamning.
Logikken her er imidlertid snudd om. Logisk ville det være om felles avstamning var sann, at en kunne konstruere ett unikt fylogenetisk tre. Det beste en kan si ved motsatt logikk, er at et unikt fylogenetisk tre ville være konsistent med felles avstamning. Når alt kommer til alt, er et godt fylogenetisk tre også konsistent med skapelse. Levende organismer ble først satt i en logisk sammenheng av den svenske naturalisten C. Linne (1707-1778), som i likhet med alle vitenskapsmenn på den tiden arbeidet under forutsetning av skapelse. Hans klassifikasjonssystem er enda i bruk. Darwin avviser, uten forklaring, sammenhengen mellom klassifikasjons-systemet og en Skapelse -som søkt. Som vi skal se, fortjener overensstemmelsen med felles avstamning bedre den etiketten.
Selv om fylogenetiske (slektskaps) trær tradisjonelt er blitt konstruert ved å sammenlikne anatomiske trekk, så er det i de siste tiårene blitt populært å gjøre det ved å sammenlikne aminosyre-sekvenser til proteiner eller DNA-nukleotid sekvenser. Organismer som deler ett felles protein, som typisk er en kjede på noen få hundre aminosyrer, ville bli antatt å ha en siste felles stamfar som var den første med det proteinet. Dawkins argumenterer for hvorfor slike fylogenetiske trær er bevis for evolusjon, ved at et slikt tre ikke er hva du ville forvente om en Skaper med oversikt over hele dyreriket hadde plukket ut de 'beste proteinene til jobben.' Det virker litt absurd når Dawkins, erke-ateisten, benytter et teologisk argument for å støtte dogmet om evolusjon. Imidlertid feiler hans argument for evolusjon, ikke bare fordi det hviler på amatør-teologi, men også fordi det faktisk er feilaktig.
Et argument for felles avstamning ville bli hjulpet om anatomiske data og molekylære data alltid førte til samme treet. Men det gjør de ikke (Heled og Drummond, 2010, Rosenberg og Degnan (2010), Nichols 2001). Fylogenetiske trær basert på ulike gener er kjent for å gi motsigende resultater. Det var håp om at bruk av hele genom, eller i det minste store porsjoner av genomet, ville løse selvmotsigelsene for fylogenetiske studier. Men det gjorde bare problemet verre (Jeffroy et al. 2006).
Mangel på unike fylogenetiske (slektskaps-) trær, blir ofte bortforklart ved såkalt 'konvergent evolusjon'. Det er forekomsten av samme trekk eller karakter i uavhengige slektslinjer. Imidlertid er det en oppfinnelse, som ble funnet opp for å unngå å adressere det mislykkede ved fylogenetiske trær til å støtte felles avstamning. Det er ikke noe teoretisk støtte for 'konvergent evolusjon', og det som har vært levert, er produkt av sirkelargumentasjon. R. Dawkins (2009) synes å more seg i å beskrive atskillige eks. på 'konvergent evolusjon' uten å innse at hvilke som helst av disse ødelegger hans sak for evolusjon (-se under).
Konvergent evolusjon er blitt påkalt over hele dyre og plante-riket. Et fascinerende eks. på konvergent evolusjon er nylig rapportert mellom hørselssystemer til mammuter og insekter (Montealegre-Z et al. 2012). Øret hos virveldyr konverterer lydbølger til elektriske nerve-signaler, som går til hjernen. Lydbølger i lufta når øret, går gjennom øre-kanalen og treffer øre-trommehinnen. Trommehinnen er knyttet til et hørsels-apparat bestående av hammer, ambolt og stigbøyle i mellomøret -hos mennesker. Svingninger i øre-membranen overføres til ørevæske i sneglehuset. Den tydelige kompleksiteten i sammenkjedingen av disse øreknoklene er designet for å tilpasse den effektive motstanden (impedansen) i luft til den i ørevæske i sneglehuset. Slik forhindres refleksjoner fra grensesnittet og påfølgende signaltap. Vibrasjoner i ørevæsken blir i sin tur overført til en samling akustiske detektorer i hårceller nedsenket i væsken. Disse omgjør lydsignalene i sine frekvens-komponenter. Vibrasjoner i hårceller blir omgjort til elektriske nerve-signaler av cellene knyttet til hvert hår, og disse signalene overføres så til hjernen.
Bilde 3 og 4. Eks. på 'konvergent evolusjon' av hørsel i uavhengige slektslinjer (mammut og løvgresshoppe)
Den sør-amerikanske løvgresshoppa (Copiphora gorgonensis) er blitt oppdaget å ha et hørsels-system analogt til det for virveldyr. Den har: en dekkende membran, analog til trommehinnen,-et overføringssystem med komponenter til å overføre vibrasjonene til væske og -for matching av impedans (motstand), analogt til de tre øreknoklene i øret hos virveldyr, og -en akustisk blære fylt med væske analog til sneglehuset, som overfører lydsignaler til en frekvens-analyserende enhet, som sender analyserte signaler til hjernen.
Forfatterne kaller dette et bemerkelsesverdig tilfelle av 'konvergent evolusjon', hvor mammuter og løvgresshopper 'har utviklet seg til å høre på en bemerkelsesverdig analog måte.' Forfatteren (Dr. Lee Spetner 1997) har vist at evolusjon ikke kan fungere i henhold til det neo-darwinistiske dogmet, om det ikke ved hvert stadium av evolusjon er mange potensielt tilpasningsdyktige mutasjoner, der tilfeldighet kunne virke (noe som ingen har vist at det kan være). Men om det er, så kan det knapt være noen 'konvergent evolusjon', fordi antallet av mulige evolusjonære stier fra ethvert stadium, ville være enormt stort, noe som gjør en repetisjon av tidligere steg høyst usannsynlig, og neo-darwinistisk evolusjon dermed uforutsigbar (For øvrig er det uenighet mellom darwinister hvorvidt og i hvilken utstrekning tilpasset evolusjon er forutsigbar -Wood et al. 2005). Om det er tilstrekkelig antall tilpasningsdyktige mutasjoner i hvert steg til å tillate neo-darwinistisk evolusjon å virke, så må 'konvergent evolusjon' {i praksis-oversetters tilføyelse} være umulig.
Den innebygde motsigelsen mellom 'konvergent evolusjon' og evnen til neo-darwinistisk evolusjon å virke i det hele, setter evolusjonsteorien 'mellom en klippe og et objekt av hard materie'. Forfatteren tror disse vurderingene har inspirert St. Gould (1989) til å erklære, med hensyn til sitt hypotetiske eksperiment ved å 'gjenspille livets tape': "Jeg tror at .. hvilken som helst gjenspilling av tapen ville lede evolusjonen ned en gangvei, radikalt ulik den veien som faktisk ble tatt."
Fordi den tilfeldige naturen i neo-darwinistisk teori krever at hvilken som helst 'avspilling av tapen' ikke vil gi samme resultat en annen gang, så vil hvilken som helst konstruksjon av livets tre heller ikke ha like karakteristika -som dukker opp uavhengig, på separate grener. Det skulle ikke forefinnes noen konvergent evolusjon. I følge teorien, skulle alle sett av felles karakteristika være sporbare til en unik stamfar. Men det viser seg at fylogenetiske trær ikke fullfører denne betingelsen. Selv om Goulds syn er fast basert på neo-darwinistisk paradigme, så viste det seg ikke å holde stikk da det ble tetet eksperimentelt. (Losos et al. 1998, Blount et al. 2008). Det er i seg selv et tegn på mistak i den neo-darwinistiske teorien. Det er mange eks. i klassifikasjonssystemet til levende organismer, som ødelegger betingelsen at det ikke skulle være noen eks. på organismer som har liknende komplekse karakteristika, som ikke er sporbare til en unik stamfar. Slike forhold burde ha medført at neo-darwinister sluttet å benytte fylogenetisk tre som bevis for felles avstamning. Neo-darwinister blir tvunget til å trekke fram konvergent evolusjon i nesten samtlige diskusjoner om evolusjon:
Andre interessante eks. på 'konvergens' som ikke kan redegjøres for av nåværende evolusjonære teorier er:
Hvaler, delfiner og flaggermus er de eneste pattedyr som har ekkolokaliserings-system. Disse systemene tillater dyrene å 'se' ved hjelp av lydbølger, som i sonar-systemer (Living waters-trailer). Dyrene sender ut lydsignaler og lokaliserer objekter i sine omgivelser, ved å analysere ekkoet i retursignalene. De må være i stand til å sende sterke lydsignaler og være i stand til å oppdage svake retur-signaler fra objekter i omgivelsene. Dyrenes hjerne må være i stand til å tolke disse signalene, liksom våre hjerner fortolker synlige input fra våre øyne. På grunnlag av sin kroppsbygning er hvaler og delfiner klassifisert nærmere kua og andre drøvtyggere, enn til flaggermus. Men ut fra ekko-lokaliseringssystemet, burde de grupperes i lag med flaggermus. Liu et al (2010) har erklært: "Evnen til noen flaggermus og tann-hvaler til å produsere sonare pulser og behandle retur-ekkoet for oppdagelse av bytte og lokalisering, er et spektakulært eks. på fenotype-konvergens i pattedyr." En gruppering ut fra molekylær analyse av proteinet 'prestin', fører til samlokalisering av hvaler og delfiner i lag med flaggermus. Prestin er et lyd-sensitivt protein som er viktig for hørsel generelt, og spesielt er det del av hørselsmekanismen i forbindelse med ekko-lokaliserings systemet. Prestin-molekyler i hvaler og delfiner har 14 amino-syrer som ikke er i prestin-molekylet i noe annet pattedyr enn flaggermus Liu et al (2010). Dette er sterkt bevis for at det fylogenetiske (slektskaps) tre ikke representerer objektiv realitet, og at det skulle legges et likklede over begrepet 'felles avstamning'. Men darwinister 'redder' felles avstamning ved å erklære at prestin-proteinet og hele ekkolokaliserings-systemet i hvaler, delfiner og flaggermus er resultat av 'konvergent evolusjon'. Men de har som nevnt ikke vist at 'konvergent evolusjon' er noe annet enn deres eget påfunn.
Bilde 5 og 6: To flaggermus med uavhengig utviklet ekkolokaliserings-system?
Selv innenfor arten flaggermus, måtte 'konvergent evolusjon' ha oppstått separat i ulike arter. Både flaggermus med mustasje og hestesko-flaggermus har ekkolokaliserings-system som er tenkt å ha utviklet seg uavhengig/separat. Neuweiler (2003), kaller dette, i det han tydelig prøver å redde felles avstamning: 'ett av de mest slående eks. på konvergent evolusjon'. Både insekter og pattedyr oppdager kjemiske lukter i omgivelsene gjennom odør-reseptorer på overflata av cellene, i det sensoriske overflatevevet i lukteorganet. Dette organet inneholder et stort antall odør-receptor gener: ca. 60 i bananflue (Drosophila) og ca 1000 i mus (Thorne et al, 2004). I kombinasjon tillater disse reseptorene dyrene å skjelne mellom hundretusener av ulike odører (Buck og Axel, 1991). Hjernen mottar de kombinerte nevrologiske signalene fra disse cellene og fortolker korreponderende odører.
Øyet er sagt å ha utviklet seg uavhengig minst 40 ganger og antagelig så mye som 65 ganger {hos ulike arter -oversetters kommentar} (Salvini-Plawen og Mayr, 1977), sitert av Land og Fernald(1992). For å redde felles avstamning, må darwinister påkalle 'konvergent evolusjon', selv om de ikke har noen forestilling om hvordan den kan skje.
Bilde 7. Øyet skal ha utviklet seg uavhengig mellom 40 og 65 ganger
Bossuyt og Milinkovich (2000) rapporterte at deres analyse av kjerne- og mitokondrielt- DNA, viser at tunnel-frosker på Madagaskar og frosker fra India utviklet seg uavhengig. Også her hevder forskerne at froskene har 'konvergert' i sine morfologiske, fysiologiske og utviklingsmessige-karakterer.
Det samme var tilfelle for fem nylig utdødde arter av honning-spisende sangfugler, som var blitt kalt 'Hawai-honningetere'. Disse var lenge var klassifisert i lag med Austalske honningspisere (Meliphagidae), fordi de så ut og oppførte seg likt med dem. Fleischer et al (2008), analyserte DNA til museums prøveeksemplarer av disse utdødde fuglene, og konkluderte med at de ikke var Meliphagidae. Igjen ble det rapportert å være et 'spesielt slående eks. av konvergent evolusjon'.
Honning-spisende insekter slik som tussmørke-svermeren (hawkmoth) sies å ha konvergert med kolibrien i størrelse, vingeslag-frekvens og stoffskifte-rate, så vel som i morfologiske, fysiologiske og biokjemiske karakteristika (Welch et al, 2006). Kolibri og insekter frambringer løft i løpet av kretsende/sirklende flyvning, ved å benytte lignende aerodynamiske mekanismer (Warrick et al, 2005). Denne likheten fornekter felles avstamning, som forfatterne igjen prøver å redde ved 'konvergent evolusjon'.
Konvergens blir påkalt som et trosforsvar for evolusjonære likheter mellom proteiner i toksiner funnet i alle animalske rekker (phyla), inkludert leddyr, skalldyr og virveldyr (Fry et al, 2009).
Denne listen der darwinister tvinges til å henføre til 'konvergent evolusjon' kunne med letthet utvides. Hvert eks. er uttrykk for uvitenhet m.h.t. hvorfor dyr som klassifiseres langt fra hverandre i fylogenetiske (slektskaps) trær, har meget komplekse sammenfallende trekk. Dilemmaet stammer fra 'darwinisme-i-hullene'{oversetters uttrykk} ut fra paradigmet om felles avstamning.
Darwinister har også en annen måte å bortforklare mangel på unike fylogenetiske trær, unntatt 'konvergent evolusjon'. De postulerer at planter og dyr kan overføre DNA-fragmenter mellom arter, en prosess som kalles 'Horisontal Gen-Overføring' (HGO). HGO er kjent å trekke inn mellom prokaryote celler, slik som bakterier. Bakterielle celler har ingen kjerne til å holde sitt DNA atskilt fra resten av cellen. De kan slynge ut DNA-fragmenter fra en celle til en annen i en form for seksuell overføring eller ved at en bakterie ganske enkelt utskiller et stykke DNA, i form av en genetisk struktur (plasmide), inn i et eksternt medium, som kan bli plukket opp av en annen bakterie. Mye bakterie-resistens vunnet ved patogene bakterier, kommer fra HGT gjennom resistente bakterier.
Det er imidlertid ingen direkte bevis av slike overføringer mellom ulike arter av eukaryote celler (med kjerne). Jan O. Andersson ved Uppsala Universitet har pekt på at det vi kjenner om gen-overføring eukaryote celler, er basert på uformelle/anekdotiske bevis (Andersson, 2009) Videre i følge Keeling og Palmer (2008), så er eks. med HGO vanligvis identifisert via ulikheter i slektskapstreet mellom arter, ut fra anatomiske og genetiske data. Dermed blir det å forklare ulikheter i fylogenetiske trær ut fra HGO et sirkulært argument. Syvanen, (2012) har uttrykt, uten referanse for øvrig, at HGO er blitt observert i laboratoriet, men ikke så langt i naturlige omgivelser. Dermed har vi ingen direkte bevis på HGO i celler med kjerne. Det er kanskje derfor darwinister foretrekker å forklare problemene ved 'slektskapstreet' ut fra 'konvergent evolusjon'. Om HGO noensinne skulle vise seg å være en viktig faktor i forbindelse med slektskapstrær, så ville hele affæren bli kastet ut i kaos (Syvanen, 2012). I ethvert tilfelle støtter ikke status ved fylogenetiske (slektskaps) -trær oppunder doktrinen om felles avstamning.
Allan Rogers (2011) skrev at transposoner er brukt til å estimere fylogeni, og at de vitner om sannhet ved felles avstamning. Han argumenterer at' siden transposoner ikke er funksjonelle og setter seg selv tilfeldig inn i genomet, så kan de bli brukt til å etablere et fylogenetisk tre'. Han skriver (s27): 'det er ekstremt usannsynlig at to transposoner noen sinne vil sette seg inn på samme sted. Det innebærer at to individer som deler samme transposon også må ha samme stamfar.'
Bilde 9. Transposoner -funksjonelle og på foretrukne steder
Det viser seg imidlertid at dette premisset er usant: 1) Transposoner er funksjonelle (Emera og Wagner, 2012). Vi har tidligere sett at de kan være ansvarlige for å reorganisere genomet, som et tilpasset svar på input fra omgivelsene.
2) De setter seg ikke tilfeldig inn. J. Shaphiro som har studert transposoner i mer enn 35 år, rapporterer at transposoner setter seg inn på foretrukne steder (Shaphiro, 2011). Det samme har andre eksperter på transposoner gjort (Levy et al. 2010). Som en konsekvens er ikke to transposoner på samme sted i genomet, noen indikasjon på at de deler en stamfar. Så mye for Roger's forsøk på å kreve bevis for felles avstamning fra transposoner.
 Her er en typisk darwinistisk påstand: "Det som evolusjon har, er hva hvilken som helst god vitenskapelig påstand har: bevis og mye av det. Evolusjon er støttet av et bredt spekter av observasjoner, gjennom feltene genetikk, anatomi, økologi, dyreatferd, paleontologi og andre. Om du ønsker å utfordre evolusjonsteorien, må du adressere de bevis som finnes. Du må vise at bevis for den enten er feilaktig eller irrelevant, eller passer bedre til en annen teori. For å gjøre dette, må en kjenne både teorien og bevisene." (talkorigins.org/faqs/faq-misconceptions.html)
Her er en typisk darwinistisk påstand: "Det som evolusjon har, er hva hvilken som helst god vitenskapelig påstand har: bevis og mye av det. Evolusjon er støttet av et bredt spekter av observasjoner, gjennom feltene genetikk, anatomi, økologi, dyreatferd, paleontologi og andre. Om du ønsker å utfordre evolusjonsteorien, må du adressere de bevis som finnes. Du må vise at bevis for den enten er feilaktig eller irrelevant, eller passer bedre til en annen teori. For å gjøre dette, må en kjenne både teorien og bevisene." (talkorigins.org/faqs/faq-misconceptions.html)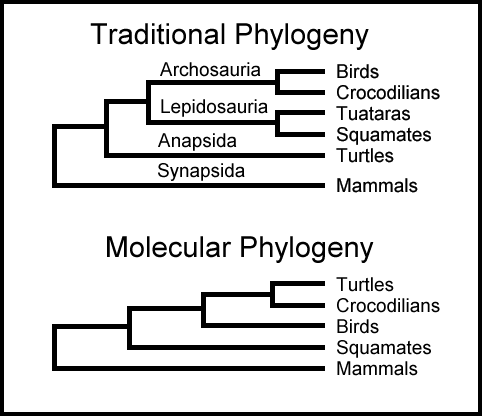 Det fylogenetiske tre som bevis for felles avstamning
Det fylogenetiske tre som bevis for felles avstamning Konvergent evolusjon er blitt påkalt over hele dyre og plante-riket. Et fascinerende eks. på konvergent evolusjon er nylig rapportert mellom hørselssystemer til mammuter og insekter (
Konvergent evolusjon er blitt påkalt over hele dyre og plante-riket. Et fascinerende eks. på konvergent evolusjon er nylig rapportert mellom hørselssystemer til mammuter og insekter ( Bilde 3 og 4. Eks. på 'konvergent evolusjon' av hørsel i uavhengige slektslinjer (
Bilde 3 og 4. Eks. på 'konvergent evolusjon' av hørsel i uavhengige slektslinjer (
 eks. på fenotype-konvergens i pattedyr." En gruppering ut fra molekylær analyse av proteinet '
eks. på fenotype-konvergens i pattedyr." En gruppering ut fra molekylær analyse av proteinet ' Øyet er sagt å ha utviklet seg uavhengig minst 40 ganger og antagelig så mye som 65 ganger {hos ulike arter -oversetters kommentar} (
Øyet er sagt å ha utviklet seg uavhengig minst 40 ganger og antagelig så mye som 65 ganger {hos ulike arter -oversetters kommentar} ( Darwinister har også en annen måte å bortforklare mangel på unike fylogenetiske trær, unntatt 'konvergent evolusjon'. De postulerer at planter og dyr kan overføre DNA-fragmenter mellom arter, en prosess som kalles 'Horisontal Gen-Overføring' (HGO). HGO er kjent å trekke inn mellom
Darwinister har også en annen måte å bortforklare mangel på unike fylogenetiske trær, unntatt 'konvergent evolusjon'. De postulerer at planter og dyr kan overføre DNA-fragmenter mellom arter, en prosess som kalles 'Horisontal Gen-Overføring' (HGO). HGO er kjent å trekke inn mellom