 Det er for øvrig funnet mange fossiler som har apelignende trekk og mange er menneskelignende. Det kan alle paleoantropologer være enige om. Når det gjelder rekonstruksjon av hele individer, for ikke å snakke om menneskets evolusjonshistorie, er det vanskelig å finne enighet. F.eks. er en uenige om hvor mange arter som har eksistert av menneskelignende aper eller apelignende mennesker. Selv om en ble enige om det, er spørsmålet om hvorvidt de er forfedre til nåtidens mennesker eller utdødde sidegrener på evolusjonstreet. En har 'Out of Africe' fløyen og 'Multiregional' fløyen, hvor de første hevder Afrika var kontinentet det skjedde, mens de andre argumenterer for at vår art har uviklet seg parallelt flere steder.
Det er for øvrig funnet mange fossiler som har apelignende trekk og mange er menneskelignende. Det kan alle paleoantropologer være enige om. Når det gjelder rekonstruksjon av hele individer, for ikke å snakke om menneskets evolusjonshistorie, er det vanskelig å finne enighet. F.eks. er en uenige om hvor mange arter som har eksistert av menneskelignende aper eller apelignende mennesker. Selv om en ble enige om det, er spørsmålet om hvorvidt de er forfedre til nåtidens mennesker eller utdødde sidegrener på evolusjonstreet. En har 'Out of Africe' fløyen og 'Multiregional' fløyen, hvor de første hevder Afrika var kontinentet det skjedde, mens de andre argumenterer for at vår art har uviklet seg parallelt flere steder.I 1996 skrev evolusjonsbiolog ved Berkely, F. Clark Howell: 'Det finnes ingen omfattende teori om den menneskelige evolusjon.. Området karkakteriseres av 'narrative behandlinger', basert kun på tynt forskningsmateriale. Det er sannsynligvis sant at et omfattende scenario om menneskets evolusjon ligger utenfor vår forståelsesramme nå og kanskje for alltid'. Geoffrey Clark, ved Arizona State University, hevder at forklaringsmodellene for menneskets evolusjon ikke er særlig annet enn et korthus. Fjerner en et kort, raser hele huset.
 sterkt sminkede skuespillere. Føy så til noen 'Hvordan' historier, om å lære å bruke redskaper etc og framstillingen er fullendt. Men disse framstillingene nevner ingenting om at fossiler plasseres inn i allerede eksisterende narrative strukturer, eller at mye hviler på formodninger og subjektivt skjønn. Aldri før i vitenskapens historie har så mange basert så mye på så lite.
sterkt sminkede skuespillere. Føy så til noen 'Hvordan' historier, om å lære å bruke redskaper etc og framstillingen er fullendt. Men disse framstillingene nevner ingenting om at fossiler plasseres inn i allerede eksisterende narrative strukturer, eller at mye hviler på formodninger og subjektivt skjønn. Aldri før i vitenskapens historie har så mange basert så mye på så lite. Beretningene om at mennesket ikke er annet enn dyr, har eksistert lenge før fortellingene ble fylt ut med det tynne forskningsmaterialet en har pr. i dag. Det ultimate ikon er bare gammeldags materialistisk filosofi, forkledd som moderne empirisk vitenskap. Siden 1970 er ikonet i stigende grad blitt brukt til å fremme læresetningen at evolusjonen ikke er retningsbestemt, og at vår eksistens er et resultat av den rene tilfeldighet.
Beretningene om at mennesket ikke er annet enn dyr, har eksistert lenge før fortellingene ble fylt ut med det tynne forskningsmaterialet en har pr. i dag. Det ultimate ikon er bare gammeldags materialistisk filosofi, forkledd som moderne empirisk vitenskap. Siden 1970 er ikonet i stigende grad blitt brukt til å fremme læresetningen at evolusjonen ikke er retningsbestemt, og at vår eksistens er et resultat av den rene tilfeldighet. Ad 1) En har i dag funnet tilstrekkelig gode sediment-bergarter fra sent PreKambrium og Kambrium til å overbevise paleontologene om at hvis det hadde fantes noen forfedre -og de hadde dannet fossiler, så ville de vært oppdaget nå. I følge James Valentine og Douglas Erwin er det slik at 'de serier av bergarter fra Kambrium vi har, er like så komplette og oppbygget under samme sedimentære forhold som lagserier som ligger er tettere på nåtiden. Allikevel er det slik at forfedre er "ukjente eller ubekreftede" for alle rekker eller klasser som dukket opp i den kambriske eksplosjon. "Den kambriske eksplosjon er ..for omfattende til at den kan bortforklares med defekter i den fossile lagserie." Flere ulike undersøkelser av den fossile lagserie fra Kambrium og opp til nå har bekreftet dette synet. I februar år 2000 når de britiske geologer M.J.Benton, M.A.Wills og R.Hitchin fram til flg.
Ad 1) En har i dag funnet tilstrekkelig gode sediment-bergarter fra sent PreKambrium og Kambrium til å overbevise paleontologene om at hvis det hadde fantes noen forfedre -og de hadde dannet fossiler, så ville de vært oppdaget nå. I følge James Valentine og Douglas Erwin er det slik at 'de serier av bergarter fra Kambrium vi har, er like så komplette og oppbygget under samme sedimentære forhold som lagserier som ligger er tettere på nåtiden. Allikevel er det slik at forfedre er "ukjente eller ubekreftede" for alle rekker eller klasser som dukket opp i den kambriske eksplosjon. "Den kambriske eksplosjon er ..for omfattende til at den kan bortforklares med defekter i den fossile lagserie." Flere ulike undersøkelser av den fossile lagserie fra Kambrium og opp til nå har bekreftet dette synet. I februar år 2000 når de britiske geologer M.J.Benton, M.A.Wills og R.Hitchin fram til flg.  konklusjon: "Den fossile lagseries tidligste deler er klart ufullstendige, men en kan saktens betrakte dem som pålitelige nok til å tegne de brede linjer i livets historie.
konklusjon: "Den fossile lagseries tidligste deler er klart ufullstendige, men en kan saktens betrakte dem som pålitelige nok til å tegne de brede linjer i livets historie. Ad 3) Den tredje måte evolusjonsbiologer har prøvd demontere den kambriske eksplosjon på, er ved å hevde at molekylært forskningsmateriale fra nålevende skapninger peker i retning av en felles stamfar til dyrerekkene, hundrevis av millioner år før Kambrium. For å forstå dette argumentet og hvorfor det ikke 'holder vann', er vi nødt til å se nærmere på 'molekylær fylogenese'
Ad 3) Den tredje måte evolusjonsbiologer har prøvd demontere den kambriske eksplosjon på, er ved å hevde at molekylært forskningsmateriale fra nålevende skapninger peker i retning av en felles stamfar til dyrerekkene, hundrevis av millioner år før Kambrium. For å forstå dette argumentet og hvorfor det ikke 'holder vann', er vi nødt til å se nærmere på 'molekylær fylogenese'  hvilke aminosyrer som inngår i en organismes mange ulike proteiner. Under replikasjonen blir rekkefølgen av baser kopiert til et nytt DNA-molekyl. Men molekylære tilfeldigheter eller mutasjoner kan en sjelden gang gjøre kopien ulik modermolekylet. Derfor kan organismer inneholde DNA-molekyler, og dermed også proteiner, som i noen grad avviker fra forfedrenes DNA og proteiner.
hvilke aminosyrer som inngår i en organismes mange ulike proteiner. Under replikasjonen blir rekkefølgen av baser kopiert til et nytt DNA-molekyl. Men molekylære tilfeldigheter eller mutasjoner kan en sjelden gang gjøre kopien ulik modermolekylet. Derfor kan organismer inneholde DNA-molekyler, og dermed også proteiner, som i noen grad avviker fra forfedrenes DNA og proteiner. Da det ble raskere teknikker for analyse av DNA-sekvenser, ble det mer alminnelig å analysere genkodene enn proteinene, slik det tidligere var gjort. Utover DNA og proteiner, inneholder også alle organismer
Da det ble raskere teknikker for analyse av DNA-sekvenser, ble det mer alminnelig å analysere genkodene enn proteinene, slik det tidligere var gjort. Utover DNA og proteiner, inneholder også alle organismer 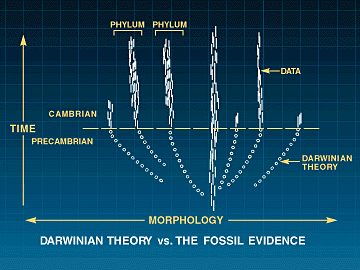 Molekylær fylogenese og den kambriske eksplosjon
Molekylær fylogenese og den kambriske eksplosjon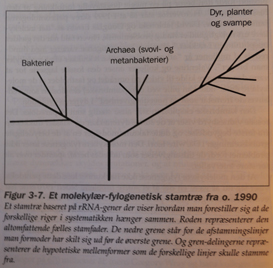 Moderne versjoner av det darwinistiske Livets stamtre, kalles "fylogenetiske (stam)tre". På et typisk slikt tre, representerer "roten" stamformen til alle andre organismer i treet. De nederste greinene tegner de avstamningslinjer som forholdsvis tidlig har skilt seg ut, mens de høyere står for senere oppdelinger. Endene av grenene viser de egentlige arter. Oppdelingspunktet mellom to grener angir den hypotetiske stamformen for de to grenene. Mange slike trær tegnes slik at grenenes lengde svarer til forskjeller i sekvenser, som i sin tur skal indikere hvor lang tid det er gått siden de gikk hver til sitt (Fig3-7)
Moderne versjoner av det darwinistiske Livets stamtre, kalles "fylogenetiske (stam)tre". På et typisk slikt tre, representerer "roten" stamformen til alle andre organismer i treet. De nederste greinene tegner de avstamningslinjer som forholdsvis tidlig har skilt seg ut, mens de høyere står for senere oppdelinger. Endene av grenene viser de egentlige arter. Oppdelingspunktet mellom to grener angir den hypotetiske stamformen for de to grenene. Mange slike trær tegnes slik at grenenes lengde svarer til forskjeller i sekvenser, som i sin tur skal indikere hvor lang tid det er gått siden de gikk hver til sitt (Fig3-7)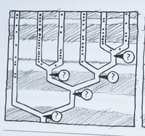 Med andre ord: ulike molekyler fører til høyst ulike fylogenetiske stamtrær. Biologen Carl Woese en av pionerene bak konstruksjonen av rRNA-baserte stamtrær, kommenterte: "Inntil videre har ingen troverdige organisme-fylogeneser sett dagens lys, ut fra de mange individuelle protein-fylogeneser en har laget. Brudd på den fylogenetiske kontinuitet forekommer overalt i stamtreet. Det mangler sammenheng mellom roten og oppover. De større forgreininger henger ikke sammen. Gruppene innbyrdes gjør det heller ikke. Og dannelsen av de grunnleggende grupperinger henger heller ikke sammen.
Med andre ord: ulike molekyler fører til høyst ulike fylogenetiske stamtrær. Biologen Carl Woese en av pionerene bak konstruksjonen av rRNA-baserte stamtrær, kommenterte: "Inntil videre har ingen troverdige organisme-fylogeneser sett dagens lys, ut fra de mange individuelle protein-fylogeneser en har laget. Brudd på den fylogenetiske kontinuitet forekommer overalt i stamtreet. Det mangler sammenheng mellom roten og oppover. De større forgreininger henger ikke sammen. Gruppene innbyrdes gjør det heller ikke. Og dannelsen av de grunnleggende grupperinger henger heller ikke sammen.  En del molekylærbiologer mener problemet er metodologisk. I følge Forterre og Phillippe forandrer noen sekvenser seg for hurtig i evolusjonsmessig henseende til å bevare en "fylogenetisk melding" over lange tidsforløp. De hevder at ved å begrense seg til sekvenser som de mener har undergått en langsom evolusjon, kan de nå fram til et allmenngyldig stamtre. Problemet deres er at deres analyser peker i retning av en celle med kjerne som den altomfattende 'felles stamfar'. Tidligere er alltid darwinister gått ut fra at en celle uten kjerne er mer opprinnelig enn celler med kjerne. Ut fra darwinistisk synspunkt har 'treet slått rot på feil sted.'
En del molekylærbiologer mener problemet er metodologisk. I følge Forterre og Phillippe forandrer noen sekvenser seg for hurtig i evolusjonsmessig henseende til å bevare en "fylogenetisk melding" over lange tidsforløp. De hevder at ved å begrense seg til sekvenser som de mener har undergått en langsom evolusjon, kan de nå fram til et allmenngyldig stamtre. Problemet deres er at deres analyser peker i retning av en celle med kjerne som den altomfattende 'felles stamfar'. Tidligere er alltid darwinister gått ut fra at en celle uten kjerne er mer opprinnelig enn celler med kjerne. Ut fra darwinistisk synspunkt har 'treet slått rot på feil sted.'  En annen løsning på problemet er framsatt av W.Ford Doolittle, biolog ved Dalousie University. Han skriver i 1999: "Det kan godt være at det ennå ikke er lykkes .. å finne det riktige tre,..fordi livets historie i bunn og grunn ikke kan framstilles som et tre." I følge Doolittle skyldes de fylogenetiske uoverensstemmelser i overveiende grad "en gjensidig genoverføring." Doolittle lanserer den idé at genoverførsel mellom bakterier og urceller seg i mellom ligger til grunn for mange av uoverensstemmelsene i molekylære fylogeneser. Det innebærer at livets tidlige historie ikke ville likne et tre, men mer ett sammenflettet kratt (Fig3-8)
En annen løsning på problemet er framsatt av W.Ford Doolittle, biolog ved Dalousie University. Han skriver i 1999: "Det kan godt være at det ennå ikke er lykkes .. å finne det riktige tre,..fordi livets historie i bunn og grunn ikke kan framstilles som et tre." I følge Doolittle skyldes de fylogenetiske uoverensstemmelser i overveiende grad "en gjensidig genoverføring." Doolittle lanserer den idé at genoverførsel mellom bakterier og urceller seg i mellom ligger til grunn for mange av uoverensstemmelsene i molekylære fylogeneser. Det innebærer at livets tidlige historie ikke ville likne et tre, men mer ett sammenflettet kratt (Fig3-8) evolusjonstreet stemmer ikke med de større trekk i det fossile og molekylære vitnesbyrd. Den kambriske eksplosjon viser at de høyeste kategorier er oppstått først, og det vender opp ned på Darwins evolusjonstre. (Se mer i "Darwins Doubt lenger ned i bloggen") Allikevel har "Livets Stamtre" stadig en dominerende plass blant evolusjonens ikoner, fordi darwinister har utropt det til å være et faktum!
evolusjonstreet stemmer ikke med de større trekk i det fossile og molekylære vitnesbyrd. Den kambriske eksplosjon viser at de høyeste kategorier er oppstått først, og det vender opp ned på Darwins evolusjonstre. (Se mer i "Darwins Doubt lenger ned i bloggen") Allikevel har "Livets Stamtre" stadig en dominerende plass blant evolusjonens ikoner, fordi darwinister har utropt det til å være et faktum!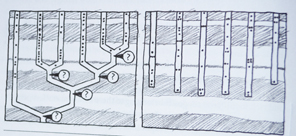 oppstått mer enn én gang og på ulike steder til ulike tider." Etter å ha fått tilgang til nyeste informasjon om molekylær fylogenese, skriver biologen Malcolm Gordon i 1998: "livet ser ut til å ha hatt atskillige opprinnelseskilder. Det universelle livets stamtre synes å ha mer enn bare en rot.. Den tradisjonelle versjon av teorien om felles avstamning, gjelder tilsynelatende ikke for riker ..[eller] rekker, og muligens heller ikke for mange av klassene innenfor rekkene.
oppstått mer enn én gang og på ulike steder til ulike tider." Etter å ha fått tilgang til nyeste informasjon om molekylær fylogenese, skriver biologen Malcolm Gordon i 1998: "livet ser ut til å ha hatt atskillige opprinnelseskilder. Det universelle livets stamtre synes å ha mer enn bare en rot.. Den tradisjonelle versjon av teorien om felles avstamning, gjelder tilsynelatende ikke for riker ..[eller] rekker, og muligens heller ikke for mange av klassene innenfor rekkene. Da en kinesisk paleontolog, Jun-Yuan Chen, kom
Da en kinesisk paleontolog, Jun-Yuan Chen, kom 
 Mikrotuber: rørformede, 24 nm tykke fibrer, bygd opp av proteinene α- og ß-tubulin. Slike fibrer brukes ved celledelingen , ved intracellulær transport (se kinesin ) og i flimmerhår. (snl)
Mikrotuber: rørformede, 24 nm tykke fibrer, bygd opp av proteinene α- og ß-tubulin. Slike fibrer brukes ved celledelingen , ved intracellulær transport (se kinesin ) og i flimmerhår. (snl)
 Ny kunnskap om arvematerialet
Ny kunnskap om arvematerialet data-filer, tilgjengelig for gjenfinning og kontekst-avhengig uttrykk ved et komplekst informasjons-prosesserings system. Det tilsvarer mer hvordan data lagres på en harddisk, med pekere og der det er fysisk mulig og med mest mulig nærhet. Cellens informasjonssystem inneholder således kjennetegn vi hittil bare har funnet i intelligent konstruerte systemer. Basert på vår nåværende kjennskap til årsak og virknings-lovene i verden, kjenner vi bare én mulig årsak til slike systemer -intelligens. At ID slik kan forstolke ny informasjon, understreker dets evne til å gi et fortolkende rammeverk for biologisk forskning.
data-filer, tilgjengelig for gjenfinning og kontekst-avhengig uttrykk ved et komplekst informasjons-prosesserings system. Det tilsvarer mer hvordan data lagres på en harddisk, med pekere og der det er fysisk mulig og med mest mulig nærhet. Cellens informasjonssystem inneholder således kjennetegn vi hittil bare har funnet i intelligent konstruerte systemer. Basert på vår nåværende kjennskap til årsak og virknings-lovene i verden, kjenner vi bare én mulig årsak til slike systemer -intelligens. At ID slik kan forstolke ny informasjon, understreker dets evne til å gi et fortolkende rammeverk for biologisk forskning.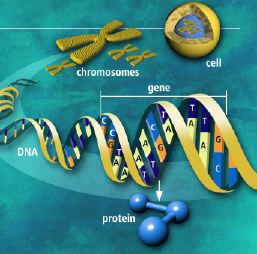 Det er tre oppdagelser angående cellens informasjonssystem som illustrerer ID's evne til å forklare ellers vanskelig forståelige oppdagelser: 1. Funksjonell, spesifikk informasjon er tett konsentrert i DNAet. 2) Genomet (en celles totale mengde av genetisk materiale, dvs. samtlige gener på alle kromosomer) er
Det er tre oppdagelser angående cellens informasjonssystem som illustrerer ID's evne til å forklare ellers vanskelig forståelige oppdagelser: 1. Funksjonell, spesifikk informasjon er tett konsentrert i DNAet. 2) Genomet (en celles totale mengde av genetisk materiale, dvs. samtlige gener på alle kromosomer) er  Mens evolusjonister før proklamerte at mesteparten av DNAet var søppel (junk-DNA), har en måttet endre oppfatning av dette. Genetisk informasjon i DNA er organisert for å maksimere informasjonstettheten. Selv om noe innhold er degradert over tid, så er den store majoriteten av base-sekvenser i genomet essensielle for å utøve biologiske funksjoner. Dette gjelder også de mange sekvensene som ikke er koder for protein. Genetiske signaler er altoverveiende i forhold til 'støy', slik ID-tilhengere forutsa tidlig på 1990-tallet.
Mens evolusjonister før proklamerte at mesteparten av DNAet var søppel (junk-DNA), har en måttet endre oppfatning av dette. Genetisk informasjon i DNA er organisert for å maksimere informasjonstettheten. Selv om noe innhold er degradert over tid, så er den store majoriteten av base-sekvenser i genomet essensielle for å utøve biologiske funksjoner. Dette gjelder også de mange sekvensene som ikke er koder for protein. Genetiske signaler er altoverveiende i forhold til 'støy', slik ID-tilhengere forutsa tidlig på 1990-tallet. Genom studier viser også at cellen adresserer distribuerte genetiske data-sett og så henter spesifikk informasjon for å styre produksjonen av protein, mye lik operativisystemet i en computer. Mer enn enkle strenger, blir det snakk om spesifiserte base-selvenser plassert rundt om på DNA-spiralen, noen ganger også rundt på ulike kromosomer. I tillegg til genetisk kode, bruker cellens informasjonssystem andre høy-nivå koder som bestemmer hvordan ulike moduler av genetisk informasjon skal hentes og behandles før oversettelse finner sted.
Genom studier viser også at cellen adresserer distribuerte genetiske data-sett og så henter spesifikk informasjon for å styre produksjonen av protein, mye lik operativisystemet i en computer. Mer enn enkle strenger, blir det snakk om spesifiserte base-selvenser plassert rundt om på DNA-spiralen, noen ganger også rundt på ulike kromosomer. I tillegg til genetisk kode, bruker cellens informasjonssystem andre høy-nivå koder som bestemmer hvordan ulike moduler av genetisk informasjon skal hentes og behandles før oversettelse finner sted. DNA-sekvenser (introns) som tidligere ble ansett for å være 'meningsløse' er nå funnet å romme gener som kan bygge proteiner. I tillegg har individuelle introns implantert i seg koder for regulerende og strukturelle RNA-er. Slik sett opptrer de som russiske dukker, som inneholder multiple beskjeder inn i seg selv, og selv er del av en større genetisk beskjed. (Fra dataspråket kjenner vi Object-Linked-Embedding (OLE), som gjør at et objekt skapt av én applikasjon kan innlemmes i et dokument skapt av en annen applikasjon, uten å miste noen kjennetegn..)
DNA-sekvenser (introns) som tidligere ble ansett for å være 'meningsløse' er nå funnet å romme gener som kan bygge proteiner. I tillegg har individuelle introns implantert i seg koder for regulerende og strukturelle RNA-er. Slik sett opptrer de som russiske dukker, som inneholder multiple beskjeder inn i seg selv, og selv er del av en større genetisk beskjed. (Fra dataspråket kjenner vi Object-Linked-Embedding (OLE), som gjør at et objekt skapt av én applikasjon kan innlemmes i et dokument skapt av en annen applikasjon, uten å miste noen kjennetegn..) Kombinasjon av gen-supermapper, grupperes inn i enda større sammenhenger av 'like-gjøremål' (isochores). Disse isochores har som oppgave å regulere relasjoner mellom nærliggende kromosomer i cellekjernen. De tjener også som et rammeverk for dannelse av organell-liknende samlinger. Dette er trolig bare de lavere lagene av ennå uoppdagede høyere lags koder. Datafolk får en slags déja-vu når vi møter hierarkiske fil-strukturer i biologiske organismer.
Kombinasjon av gen-supermapper, grupperes inn i enda større sammenhenger av 'like-gjøremål' (isochores). Disse isochores har som oppgave å regulere relasjoner mellom nærliggende kromosomer i cellekjernen. De tjener også som et rammeverk for dannelse av organell-liknende samlinger. Dette er trolig bare de lavere lagene av ennå uoppdagede høyere lags koder. Datafolk får en slags déja-vu når vi møter hierarkiske fil-strukturer i biologiske organismer.  I de siste årene har utviklings og evolusjons-biologer oppdaget at kodende sekvenser i genomet ikke alene bestemmer funksjonen til genproduktet under embryo-utviklingen. Ofte er det den større konteksten(sammenhengen) genene finnes i, som bestemmer den spesifikke funksjonen til proteinene de utvikler. ved å sammenligne gener mellom grupper, har biologer oppdaget at hovedsakelig identiske sekvenser regulerer meget forskjellige strukturer i ulike organismer.
I de siste årene har utviklings og evolusjons-biologer oppdaget at kodende sekvenser i genomet ikke alene bestemmer funksjonen til genproduktet under embryo-utviklingen. Ofte er det den større konteksten(sammenhengen) genene finnes i, som bestemmer den spesifikke funksjonen til proteinene de utvikler. ved å sammenligne gener mellom grupper, har biologer oppdaget at hovedsakelig identiske sekvenser regulerer meget forskjellige strukturer i ulike organismer. gener kontrollerer utviklingen og at like gener skulle danne like organismer og strukturer. Denne oppdagelsen problematiserer også påstander om likt opphav, når samme gentype finnes i 'fjernt beslektede' arter.
gener kontrollerer utviklingen og at like gener skulle danne like organismer og strukturer. Denne oppdagelsen problematiserer også påstander om likt opphav, når samme gentype finnes i 'fjernt beslektede' arter. Dette trigger spørsmålet: Hvordan kunne gener og proteiner ha overlevd og formert seg, før den eksterne og komplekse organisme-konteksten, som bestemmer virkemåten, fantes? Dette trekket ved gener synes forbløffende (puzzling) for evolusjonsteorien. Det blir ikke bare et super-komplisert puslespill, men å holde det i live uten 'styring ovenfra' synes uholdbart. Ut fra ID som har rom for et forsyn, en planleggende intelligens, virker imidlertid dette ikke ødeleggende.
Dette trigger spørsmålet: Hvordan kunne gener og proteiner ha overlevd og formert seg, før den eksterne og komplekse organisme-konteksten, som bestemmer virkemåten, fantes? Dette trekket ved gener synes forbløffende (puzzling) for evolusjonsteorien. Det blir ikke bare et super-komplisert puslespill, men å holde det i live uten 'styring ovenfra' synes uholdbart. Ut fra ID som har rom for et forsyn, en planleggende intelligens, virker imidlertid dette ikke ødeleggende. Biologer vet ikke hvor alle denne informasjonen utenom genomet befinner seg, men har lokalisert noe. For eks. vet de at strukturen og lokaliseringen til celle-skjelettet spiller inn på utviklingen av embryoet. I tillegg påvirker lokalisering av spesifikke mål-steder på innsiden av celle-membranen formen på celle-skjelletet og influerer utviklingen av organisme-formen. Selv om disse er laget av proteiner, er det ikke bare proteinet, men plasseringen og den tre-dimensjonale strukturelle informasjonen som bestemmer strukturen på celleskjelettet og lokaliseringen av dets mindre enheter. I encellede organismer kan kutt i celle-membranen danne arvelige endringer i membran-mønstre, selv om DNA ikke er skadet.
Biologer vet ikke hvor alle denne informasjonen utenom genomet befinner seg, men har lokalisert noe. For eks. vet de at strukturen og lokaliseringen til celle-skjelettet spiller inn på utviklingen av embryoet. I tillegg påvirker lokalisering av spesifikke mål-steder på innsiden av celle-membranen formen på celle-skjelletet og influerer utviklingen av organisme-formen. Selv om disse er laget av proteiner, er det ikke bare proteinet, men plasseringen og den tre-dimensjonale strukturelle informasjonen som bestemmer strukturen på celleskjelettet og lokaliseringen av dets mindre enheter. I encellede organismer kan kutt i celle-membranen danne arvelige endringer i membran-mønstre, selv om DNA ikke er skadet. I det hele bestemmes formen og strukturen til cella av både gener og tidligere tredimensjonale-strukturer og deres organisering. Proteiner transporteres og finner fram, til riktig adresse delvis p.g.a. tidligere tredimensjonale-strukturer og deres organisering. (Nobel-prisen i medisin 2013 gikk til celle-biologer for deres arbeid med cellens transport-system..) Gener forsyner nødvendig, men ikke tilstrekkelig informasjon for utvikling av de tre dimensjonale former og strukturer de produserer i celler og for kroppsbygning. Således kan gener mutere i det uendelige uten at de påvirker større formmessige endringer.
I det hele bestemmes formen og strukturen til cella av både gener og tidligere tredimensjonale-strukturer og deres organisering. Proteiner transporteres og finner fram, til riktig adresse delvis p.g.a. tidligere tredimensjonale-strukturer og deres organisering. (Nobel-prisen i medisin 2013 gikk til celle-biologer for deres arbeid med cellens transport-system..) Gener forsyner nødvendig, men ikke tilstrekkelig informasjon for utvikling av de tre dimensjonale former og strukturer de produserer i celler og for kroppsbygning. Således kan gener mutere i det uendelige uten at de påvirker større formmessige endringer. En analogi kan sees til fysikk, der organiseringen av transistorer, motstand og kondensatorer er det som bestemmer virkemåten av elektronikken, ikke delene i seg selv. Organismer inneholder både informasjonsrike proteiner og gener, og danner informasjonsrike arrangement av disse komponentene i form av hierarkisk oppbygde systemer. Et rikt flerlags-informasjon hierarki er ikke uventet fra ID-hold. At biologiske organismer har slik oppbygning understreker ytterligere relevansen til ID.
En analogi kan sees til fysikk, der organiseringen av transistorer, motstand og kondensatorer er det som bestemmer virkemåten av elektronikken, ikke delene i seg selv. Organismer inneholder både informasjonsrike proteiner og gener, og danner informasjonsrike arrangement av disse komponentene i form av hierarkisk oppbygde systemer. Et rikt flerlags-informasjon hierarki er ikke uventet fra ID-hold. At biologiske organismer har slik oppbygning understreker ytterligere relevansen til ID.