Saksområde: |
Påvisning: |
Begrunnelse: |
| Mønstre i cytoskjellettet |
Plassering avgjør- en form for kritisk informasjon |
Arves direkte |
| Mikrtotubuler |
Lokasjon og arrangement avgjør arv |
|
| Ione-kanaler og elektromagn. felt |
Arrangering og plassering av ione-kanaler kan påvirke dyrets utvikling |
Kunstige felt kan forårsake celle-vandring. |
| Sukkermolekyler på cellemembran |
Plassering spiller kritisk rolle i funksjonen de har |
|
| Mål-punkter på celle-membran |
Spesifikk lokasjon og fordeling avgjør funksjon |
Plassering avgjøres ikke av genprodukt alene |
| Folding av proteiner |
Mange proteiner folder seg ulikt avhengig av omgivelsene |
Påvist i sammenlignende utviklingsbiologi |
| Morfogenetiske proteiner, regulerende gener og dGRNs |
Disse avhenger alle av plasseringen av spesifikke, informasjonsrike, tidligere cellestrukturer |
En har aldri opplevd at dGRNs er blitt endret med livskraftig avkom som følge |
| Cellestruktur |
Resultat av både genetikk og tidligere 3-dim-struktur og organisering i celle, cellemembran, og cytoskjellett |
Påvist ved forsøk at snitt i cellemembran arves direkte på encellede organismer |
 Når en gjennomsnittlig norsk professor skal snakke om livets store spørsmål, og prøver gi et vitenskapelig svar på de store spørsmålene: 'hvor kommer vi fra?' 'Hvor går vi hen?' og 'Hvorfor er vi her?' I de tilfellene er det
Når en gjennomsnittlig norsk professor skal snakke om livets store spørsmål, og prøver gi et vitenskapelig svar på de store spørsmålene: 'hvor kommer vi fra?' 'Hvor går vi hen?' og 'Hvorfor er vi her?' I de tilfellene er det  samme svar som kommer tre ganger: 'Darwin med sin lære om mutasjoner og utvalg'. Menneskehetens tre eksistensielle spørsmål påkaller bare snusfornuftige skuldertrekk fra en darwinist. Svarene som en typisk ortodoks darwinist er i stand til å gi på disse tre er bare utrolig grunne og forutsigbare:
samme svar som kommer tre ganger: 'Darwin med sin lære om mutasjoner og utvalg'. Menneskehetens tre eksistensielle spørsmål påkaller bare snusfornuftige skuldertrekk fra en darwinist. Svarene som en typisk ortodoks darwinist er i stand til å gi på disse tre er bare utrolig grunne og forutsigbare: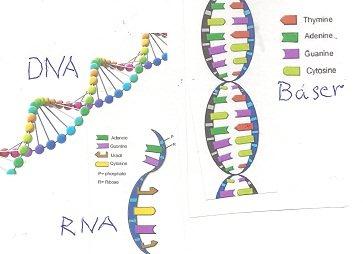 Noen av oss lærte som barn at 'alt det gode kommer ovenfra'. Grunntanken i darwinismen er motsatt: 'Alle årsaker kommer nedenfra'. I darwinistisk tankegang er ikke livet noe som skjenkes oss ovenfra, tvert i mot kommer det fra pølen der nede. Livet kommer snikende fra en uidentifiserbar amøbe, nede i 'ursuppa'. Denne amøben har dratt seg opp etter håret den ikke har, og er blitt mer og mer avansert på sin vei til å bli menneske. Det må være et filosofisk og logisk problem for darwinismen at alle forklaringer er fattigere enn fenomenene de skal forklare. Ved stadig reduksjon blir uvegerlig noe borte underveis. Det at liv spontant skal ha blitt til av ikke-liv er et slikt eksempel.
Noen av oss lærte som barn at 'alt det gode kommer ovenfra'. Grunntanken i darwinismen er motsatt: 'Alle årsaker kommer nedenfra'. I darwinistisk tankegang er ikke livet noe som skjenkes oss ovenfra, tvert i mot kommer det fra pølen der nede. Livet kommer snikende fra en uidentifiserbar amøbe, nede i 'ursuppa'. Denne amøben har dratt seg opp etter håret den ikke har, og er blitt mer og mer avansert på sin vei til å bli menneske. Det må være et filosofisk og logisk problem for darwinismen at alle forklaringer er fattigere enn fenomenene de skal forklare. Ved stadig reduksjon blir uvegerlig noe borte underveis. Det at liv spontant skal ha blitt til av ikke-liv er et slikt eksempel.  1) Tilfeldige mutasjoner får æren for oppbygningen av biologiens orden og genetiske informasjon. I virkeligheten virker mutasjonene ensidig og irreversibelt nedbrytende på biologisk orden og genetisk informasjon.
1) Tilfeldige mutasjoner får æren for oppbygningen av biologiens orden og genetiske informasjon. I virkeligheten virker mutasjonene ensidig og irreversibelt nedbrytende på biologisk orden og genetisk informasjon.  Denne observasjonen ligger helt i tråd med konklusjonene fra matematikeren Fred Hoyle. Forskerne er, i følge Behe, langt høyere grad i stand til å avgjøre hvor grensene for evolusjonen går, når en kjenner det genetiske grunnlaget for mutasjonene. Denne viten brukes på en konkret sak, som har vært nøye forsket på: 'Med historien om malaria, står vi ved den aller beste prøve på om en darwinistiske teori holder. Det skyldes malariens enorme populasjonsstørrelse, dens reproduksjonsrate og vår viten om dens genetikk. Behe påpeker at hundrevis av mutasjoner som gir en viss motstand mot malaria, er dukket opp i det menneskelige genom. Disse er spredt gjennom menneskelig populasjon gjennom naturlig seleksjon. Materialet viser imidlertid at det finnes skarpe grenser for hvilken effekt tilfeldige mutasjoner har.
Denne observasjonen ligger helt i tråd med konklusjonene fra matematikeren Fred Hoyle. Forskerne er, i følge Behe, langt høyere grad i stand til å avgjøre hvor grensene for evolusjonen går, når en kjenner det genetiske grunnlaget for mutasjonene. Denne viten brukes på en konkret sak, som har vært nøye forsket på: 'Med historien om malaria, står vi ved den aller beste prøve på om en darwinistiske teori holder. Det skyldes malariens enorme populasjonsstørrelse, dens reproduksjonsrate og vår viten om dens genetikk. Behe påpeker at hundrevis av mutasjoner som gir en viss motstand mot malaria, er dukket opp i det menneskelige genom. Disse er spredt gjennom menneskelig populasjon gjennom naturlig seleksjon. Materialet viser imidlertid at det finnes skarpe grenser for hvilken effekt tilfeldige mutasjoner har. I 1995 ble Nobel prisen i medisin gitt til to tyske forskere (Wieshaus og Nüsslein-Vollhard) for deres nye og grundige arbeid med å kartlegge genomet til bananfluer. De hadde i årtier arbeidet med mutasjonseksperimenter på disse, med nedslående resultater. De hadde nok framkalt forandringer i form av ekstremiteter, dupliseringer og endog noe funksjonsendring. Men ingenting hadde fungert i helheten. Riktignok var balanseorganer blitt endret til vinger, bare med den følge at flua mistet styringen.. Problemet var i følge Wieshaus at de trodde de hadde funnet alle genene nødvendig for å endre kroppsplanen til Drosophila (typen bananflue). Likevel er ikke resultatene lovende som råmateriale for makro-evolusjon. Det neste spørsmålet er hva som ville være de rette mutasjoner for makro-evolusjon. Og vi vet ikke svaret på det..
I 1995 ble Nobel prisen i medisin gitt til to tyske forskere (Wieshaus og Nüsslein-Vollhard) for deres nye og grundige arbeid med å kartlegge genomet til bananfluer. De hadde i årtier arbeidet med mutasjonseksperimenter på disse, med nedslående resultater. De hadde nok framkalt forandringer i form av ekstremiteter, dupliseringer og endog noe funksjonsendring. Men ingenting hadde fungert i helheten. Riktignok var balanseorganer blitt endret til vinger, bare med den følge at flua mistet styringen.. Problemet var i følge Wieshaus at de trodde de hadde funnet alle genene nødvendig for å endre kroppsplanen til Drosophila (typen bananflue). Likevel er ikke resultatene lovende som råmateriale for makro-evolusjon. Det neste spørsmålet er hva som ville være de rette mutasjoner for makro-evolusjon. Og vi vet ikke svaret på det.. Som en logisk følge av dette kan sluttes om vi treffer på trekk ved livet som hadde krevd mer enn en slik dobbelt-mutasjon, kan vi utlede at de sannsynligvis ikke er oppstått som resultat av en tilfeldig prosess i Darwins forstand. Han peker så på masser av eksempler på hvor livet er sprengfullt med slike strukturer. Han henviser f.eks. til kontrollsystemer (eller genetisk regulerende nettverk) som er involvert i oppbygning av dyrs kropper. Malaria-hiv 'eksperimentet' er gjennomført med en milliard ganger større volum enn mennesker kan utføre i laboratorier. Likevel har en ikke funnet spor av at tilfeldige mutasjoner og naturlig seleksjon, skulle ha oppbygd sammenhengende biologiske maskiner.
Som en logisk følge av dette kan sluttes om vi treffer på trekk ved livet som hadde krevd mer enn en slik dobbelt-mutasjon, kan vi utlede at de sannsynligvis ikke er oppstått som resultat av en tilfeldig prosess i Darwins forstand. Han peker så på masser av eksempler på hvor livet er sprengfullt med slike strukturer. Han henviser f.eks. til kontrollsystemer (eller genetisk regulerende nettverk) som er involvert i oppbygning av dyrs kropper. Malaria-hiv 'eksperimentet' er gjennomført med en milliard ganger større volum enn mennesker kan utføre i laboratorier. Likevel har en ikke funnet spor av at tilfeldige mutasjoner og naturlig seleksjon, skulle ha oppbygd sammenhengende biologiske maskiner.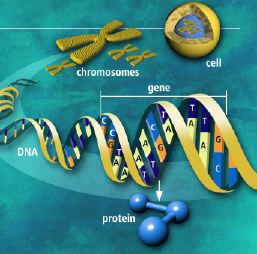 Men så må en skjelne litt: Det kommer an på øynene om ser, her også. Om det skulle være et genetisk slektskap er én ting. Å hevde at mutasjoner og naturlig utvalg er eneste mekanismer som er involvert i dette noe annet. Argumentene ovenfor, viser at det er mer enn naturlig seleksjon og mutasjoner innvolvert i dannelsen av denne genetiske nærheten. Etter oppdagelser i epigenetikken, er det f.eks. ikke 'bare' genens DNA, men også dens plassering som bestemmer dens funksjon. I følge Behe skal det noe mer enn tilfeldige mutasjoner, genetisk drift og naturlig uvalg til, og dette 'noe mer' er Input fra en designende intelligens.
Men så må en skjelne litt: Det kommer an på øynene om ser, her også. Om det skulle være et genetisk slektskap er én ting. Å hevde at mutasjoner og naturlig utvalg er eneste mekanismer som er involvert i dette noe annet. Argumentene ovenfor, viser at det er mer enn naturlig seleksjon og mutasjoner innvolvert i dannelsen av denne genetiske nærheten. Etter oppdagelser i epigenetikken, er det f.eks. ikke 'bare' genens DNA, men også dens plassering som bestemmer dens funksjon. I følge Behe skal det noe mer enn tilfeldige mutasjoner, genetisk drift og naturlig uvalg til, og dette 'noe mer' er Input fra en designende intelligens. På samme vis sier forsker i evolusjonær paleobiologi, Conway Morris, seg utilfreds med ultra-darwinsimens reduksjonistiske betraktninger. De har oppstilt et naturalistisk system, som i seg selv ikke har noe endegyldig formål, likevel lar de en viss form for hensikt (nå fram til tilnærmet samme genom) slippe inn bakdøra. Han sammenligner livet med sin evne til å navigere fram til de beste løsninger mer med en klarsynt navigatør, enn en blind urmaker. I en senere utgitt bok lyder Conway Morris sin dom over fenomenet konvergent evolusjon (evolusjon av strukturer som ligner hverandre, uten å være beslektet i darwinistisk mening), eks. vingen hos en fugl og en flaggermus: 'Ja, ettersom vår viten bryter nye grenser, vokse bare min forbløffelse'. Selv om 'den blinde urmaker' ikke vet han er på vei hjem, vet han det så likevel?
På samme vis sier forsker i evolusjonær paleobiologi, Conway Morris, seg utilfreds med ultra-darwinsimens reduksjonistiske betraktninger. De har oppstilt et naturalistisk system, som i seg selv ikke har noe endegyldig formål, likevel lar de en viss form for hensikt (nå fram til tilnærmet samme genom) slippe inn bakdøra. Han sammenligner livet med sin evne til å navigere fram til de beste løsninger mer med en klarsynt navigatør, enn en blind urmaker. I en senere utgitt bok lyder Conway Morris sin dom over fenomenet konvergent evolusjon (evolusjon av strukturer som ligner hverandre, uten å være beslektet i darwinistisk mening), eks. vingen hos en fugl og en flaggermus: 'Ja, ettersom vår viten bryter nye grenser, vokse bare min forbløffelse'. Selv om 'den blinde urmaker' ikke vet han er på vei hjem, vet han det så likevel?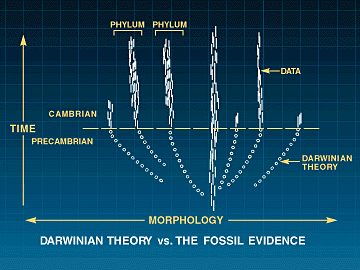 Paleontologer forstår den cambriske eksplosjonen som en plutselig geologisk tilsynekomst av nye livsformer. Å bygge disse kroppsuttrykkene krever nye utviklings program, både nye tidlig-handlende regulerende gener OG nye dGRNs. OM ingen av disse kan endres ved mutasjoner, uten å ødelegge eksisterende utviklingsprogram (og dermed dyreliv), så vil ikke naturlig utvalg ha noe å favorisere. Dermed vil utviklingen av nye dyreformer holde opp på det stadiet. Darwins tvil om fossilfunn i Cambrium er forsterket: Fossilene mangler, men i tillegg har vi et dyptgående problem med konstruksjons begrensinger. Problemet om å bygge en ny form for dyreliv, ved gradvis å endre et tett integrert system av genetiske komponenter og deres produkter infiltrert i hverandre. Ennå gjenstår mer formidable problemer for neo-Darwinismen (Kap.14-epigenetikk).
Paleontologer forstår den cambriske eksplosjonen som en plutselig geologisk tilsynekomst av nye livsformer. Å bygge disse kroppsuttrykkene krever nye utviklings program, både nye tidlig-handlende regulerende gener OG nye dGRNs. OM ingen av disse kan endres ved mutasjoner, uten å ødelegge eksisterende utviklingsprogram (og dermed dyreliv), så vil ikke naturlig utvalg ha noe å favorisere. Dermed vil utviklingen av nye dyreformer holde opp på det stadiet. Darwins tvil om fossilfunn i Cambrium er forsterket: Fossilene mangler, men i tillegg har vi et dyptgående problem med konstruksjons begrensinger. Problemet om å bygge en ny form for dyreliv, ved gradvis å endre et tett integrert system av genetiske komponenter og deres produkter infiltrert i hverandre. Ennå gjenstår mer formidable problemer for neo-Darwinismen (Kap.14-epigenetikk).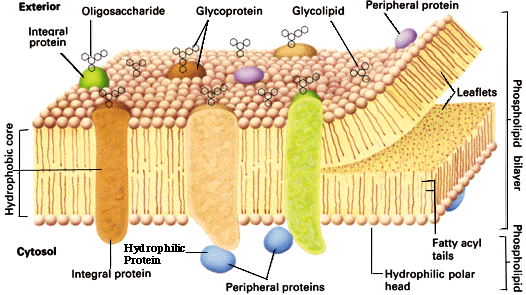
 Mens evolusjonister før proklamerte at mesteparten av DNAet var søppel (junk-DNA), har en måttet endre oppfatning av dette. Genetisk informasjon i DNA er organisert for å maksimere informasjonstettheten. Selv om noe innhold er degradert over tid, så er den store majoriteten av base-sekvenser i genomet essensielle for å utøve biologiske funksjoner. Dette gjelder også de mange sekvensene som ikke er koder for protein. Genetiske signaler er altoverveiende i forhold til 'støy', slik ID-tilhengere forutsa tidlig på 1990-tallet.
Mens evolusjonister før proklamerte at mesteparten av DNAet var søppel (junk-DNA), har en måttet endre oppfatning av dette. Genetisk informasjon i DNA er organisert for å maksimere informasjonstettheten. Selv om noe innhold er degradert over tid, så er den store majoriteten av base-sekvenser i genomet essensielle for å utøve biologiske funksjoner. Dette gjelder også de mange sekvensene som ikke er koder for protein. Genetiske signaler er altoverveiende i forhold til 'støy', slik ID-tilhengere forutsa tidlig på 1990-tallet. Dette trigger spørsmålet: Hvordan kunne gener og proteiner ha overlevd og formert seg, før den eksterne og komplekse organisme-konteksten, som bestemmer virkemåten, fantes? Dette trekket ved gener synes forbløffende (puzzling) for evolusjonsteorien. Det blir ikke bare et super-komplisert puslespill, men å holde det i live uten 'styring ovenfra' synes uholdbart. Ut fra ID som har rom for et forsyn, en planleggende intelligens, virker imidlertid dette ikke ødeleggende.
Dette trigger spørsmålet: Hvordan kunne gener og proteiner ha overlevd og formert seg, før den eksterne og komplekse organisme-konteksten, som bestemmer virkemåten, fantes? Dette trekket ved gener synes forbløffende (puzzling) for evolusjonsteorien. Det blir ikke bare et super-komplisert puslespill, men å holde det i live uten 'styring ovenfra' synes uholdbart. Ut fra ID som har rom for et forsyn, en planleggende intelligens, virker imidlertid dette ikke ødeleggende. grunnet interavhengighet av informasjonshierarkier? Gitt fenomenet fenotype- (fremtoningspreg/egenskapstype) plastisitet at organismer med samme genotype (genetisk konstitusjon/arveanlegg) har ulike fenotyper. Ut fra det kan en spørre hvor mye av variasjonen i organismer er resultat av 'preprogrammering' i motsetning til tilfeldige mutasjoner? Om gjentatte variasjoner stammer fra preprogrammering, hvor befinner den nødvendige informasjonen seg og hvordan kommer den til uttrykk?
grunnet interavhengighet av informasjonshierarkier? Gitt fenomenet fenotype- (fremtoningspreg/egenskapstype) plastisitet at organismer med samme genotype (genetisk konstitusjon/arveanlegg) har ulike fenotyper. Ut fra det kan en spørre hvor mye av variasjonen i organismer er resultat av 'preprogrammering' i motsetning til tilfeldige mutasjoner? Om gjentatte variasjoner stammer fra preprogrammering, hvor befinner den nødvendige informasjonen seg og hvordan kommer den til uttrykk?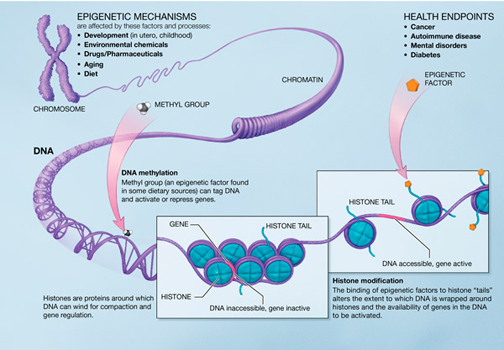
 Nå skal vi huske at 'naturlige seleksjon' her er forkortet uttrykk for neodarwinistiske syntese av mutasjon, genetisk drift og naturlige utvalg. Om vi skal se på påstandene samlet, er det første som slår oss at de beveger seg langt utover det Darwin kunne stå inne for. For påstand 1 er at naturlige seleksjon kan reduseres til et spørsmål om fysikk. Det sa aldri Darwin noe om. naturlige seleksjon går nemlig ut fra at livet allerede eksisterer, eller i det minste finnes et system som er selvreplikerende. Vi skal ta den viktige overgangen mellom ikke-levende og levende opp senere.
Nå skal vi huske at 'naturlige seleksjon' her er forkortet uttrykk for neodarwinistiske syntese av mutasjon, genetisk drift og naturlige utvalg. Om vi skal se på påstandene samlet, er det første som slår oss at de beveger seg langt utover det Darwin kunne stå inne for. For påstand 1 er at naturlige seleksjon kan reduseres til et spørsmål om fysikk. Det sa aldri Darwin noe om. naturlige seleksjon går nemlig ut fra at livet allerede eksisterer, eller i det minste finnes et system som er selvreplikerende. Vi skal ta den viktige overgangen mellom ikke-levende og levende opp senere.