Er seleksjon på genomnivå mulig?
Tradisjonelt har genetikere studert problematikk angående mutasjoner ved å betrakte én mutasjon om gangen. Så har en antatt at det som virker for én mutasjon kan utvides (ekstrapoleres) til å gjelde for alle mutasjoner. Men det er en ufornuftig holdning, á la molboenes 'tar du den, så tar du den'. Eks. 'om en har råd til én bil, så har en råd til 300 eller om en kan jonglere med 3 baller, så kan en gjøre det samme med 300'.. På samme vis ser en i genetikken at de tøffe problemene med seleksjon inntreffer når vi betrakter alle genetiske enheter kombinert i genomet. For å forstå hva som kreves for å unngå degenerasjon av genomet, må vi betrakte mutasjons og seleksjons-problematikk på genom-nivå.
Det er 3 milliarder nukleotide-posisjoner i genomet, hver med 2 kopier. Så i alt er det mulig med 6 milliarder punktmutasjoner (mulige 'trykkfeil'). Problematikken med å betrakte seleksjon av mutasjoner på genom-nivå er bare erkjent av noen få framsynte biologer i fortiden (Haldane, Muller, Kimura, Kondrashov). Igjen og igjen har hele problematikken blitt feid under teppet. Det skyldes nok ikke minst at den skaper uoverstigelige problemer for evolusjonsteorien. I de siste tiårene har vi lært at genomet er mye større og komplekst enn vi noen forutså. Vi har også lært at menneskelig mutasjonsrate er mye høyere enn en tidligere tenkte var mulig. Vi lærer at mutasjonsandelen som virkelig er nøytral, stadig synker og prosentandelen som tilfører informasjon er forsvinnende liten, om den i det hele eksisterer. Vi kan ikke lenger ignorere degenerajon i genomet, og vil se på om seleksjon på genomnivå kan løse problemet.
Konsekvensen av nevnte problem er at hver og en av oss i stigende grad er utsatt for mutasjoner. Denne destruktive mutasjons-prosessen har pågått i flere tiår. I tillegg til de ca. 100 mutasjoner vi selv legger til menneskelig genom-totalitet, så har vi arvet en hærskare av mutasjoner fra forfedrene. 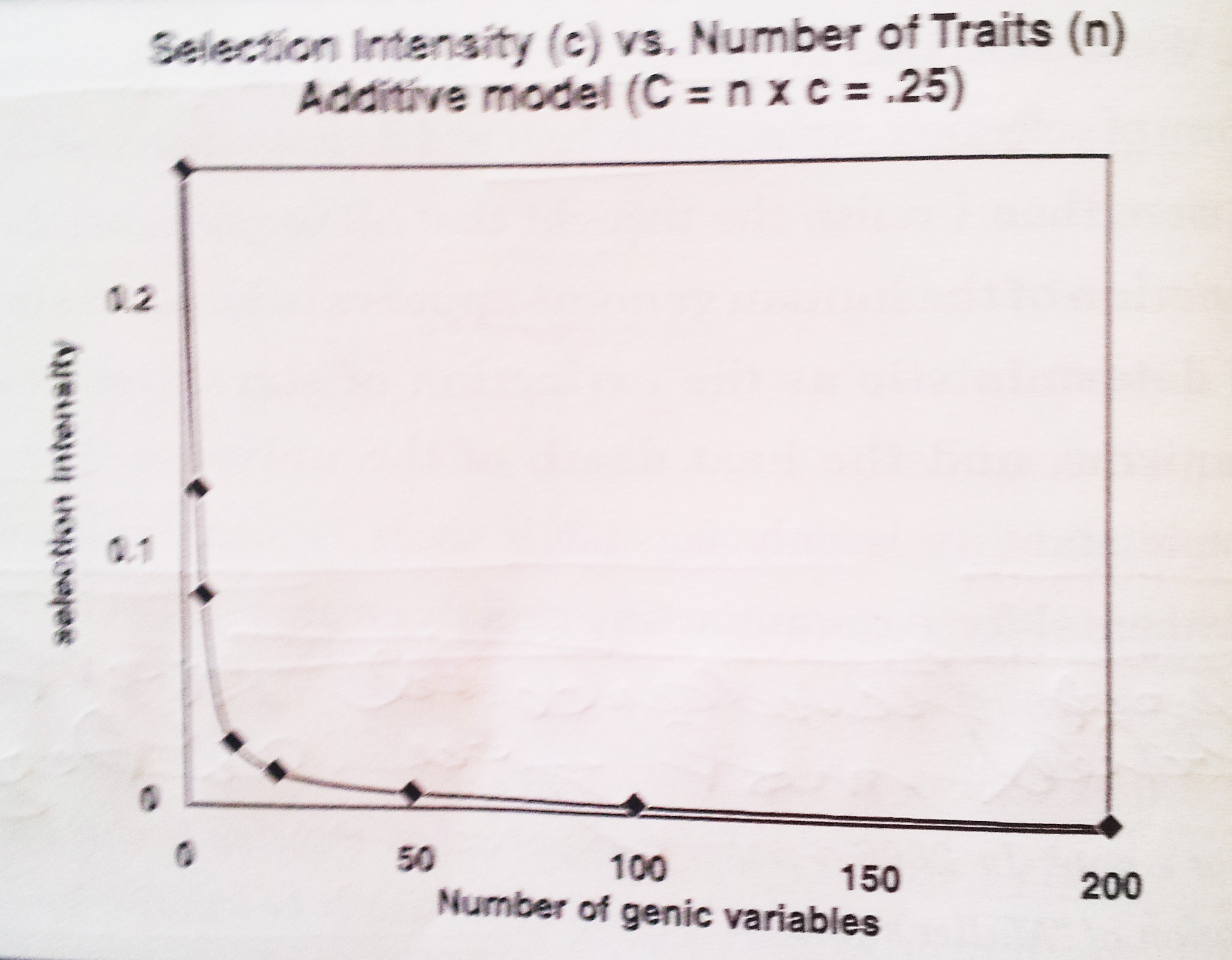 Om vi tar utgangspunkt i seleksjons-problematikk overfor enkeltmutasjoner, omtalt tidligere, så må vi multiplisere problemomfanget med en faktor på ca. 1 milliard (Tillater da at 2 milliarder nukleotider kan være 'sant nøytrale', selv om det ikke er klart at noen del av genomet kan være det.) En begynner å ane at seleksjon på genom-nivå har et helt annerledes omfang enn i forhold til seleksjon på punktmutasjoner/gennivå.
Om vi tar utgangspunkt i seleksjons-problematikk overfor enkeltmutasjoner, omtalt tidligere, så må vi multiplisere problemomfanget med en faktor på ca. 1 milliard (Tillater da at 2 milliarder nukleotider kan være 'sant nøytrale', selv om det ikke er klart at noen del av genomet kan være det.) En begynner å ane at seleksjon på genom-nivå har et helt annerledes omfang enn i forhold til seleksjon på punktmutasjoner/gennivå.
Bilde 1. Seleksjonsintensitet avtar raskt med økende antall mutasjoner (unngår for høy totalkostnad)
1. Seleksjonskostnader.
Faktumet at alle mennesker er utsatt for mutasjoner, gjør seleksjon mye vanskeligere. Hver og en av oss arver tusener av skadelige mutasjoner. Kollektivt bærer vi på hundrevis av milliarder mutasjoner, som populasjon. For å forenkle problemet kan vi se på den andelen vår generasjon har tilført menneskeheten. Om en anslagsvis regner med at hvert individ tilfører 100 mutasjon, og vi regner med en verdenspopulasjon på 6 milliarder (da boka ble skrevet), så tilfører vår generasjon 600 milliarder mutasjoner. Vi regner med en fruktbarhet på 1,5 eller at 2 mennesker tilfører 3 nye. Da har vi bare råd til å 'gi avkall på' 1 av 3, for at populasjonen skal overleve på sikt. Det er selvsagt kun teoretisk det er aktuelt, noe annet ville være å sammenlikne med nazisme ell.lign. I teorien vil en ved å selektere bort tredjeparten, likevel stå igjen med en økning på 400 milliarder mutasjoner pr. generasjon. Selv om skulle anta at to tredjedeler av disse er 'perfekt nøytrale', så har vi ennå 133 milliarder skadelige mutasjoner bare for vår generasjon. Selv om en skulle doble seleksjonsintensiteten (noe en slett ikke har råd til på sikt), har en ennå igjen 67 milliarder skadelige mutasjoner som overføres til neste generasjon. Den kjente mutasjonsraten for mennesker, er for høy til å bli møtt for noe aktuelt seleksjonsnivå. Derfor vil mutasjoner fortsette å akkumulere, og arten degenere. Når en ser det på genom-nivå, er det et forbløffende problem å forholde seg til.
2. Utydelige eller 'skjulte' mutasjoner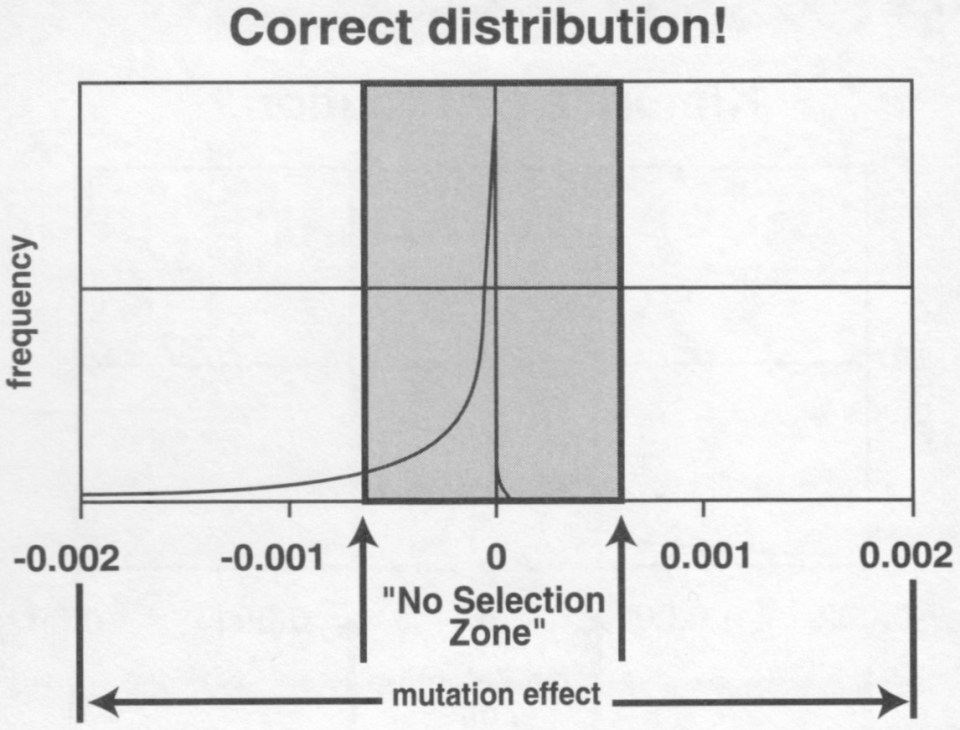
Overraskende nok er ikke dødelige eller 'nær-dødelige' mutasjoner hovedproblemer, i forhold til naturlig seleksjon. Slike problemer er sjeldne, og eliminerer gjerne seg selv. Likeledes er heller ikke 'helt-nøytrale' mutasjonsproblemer noe problem. Det er mindre mutasjoner som står for det meste av ødeleggelser, særlig i korte tidsintervall (Kimura og Ohta, 1971, s53). Seleksjon må forhindre akkumulering av mindre mutasjoner, ellers vil arten raskt forringe seg, og tilstanden ('fitness') vil avta. Selv om seleksjon kunne holde skadelige, mindre mutasjoner i sjakk, synes den maktesløs mot den mest dominerende kategorien: 'nesten nøytrale' mutasjoner. Derfor vil høyere genomer alle degenere i det lange løp, med eller uten seleksjon.
Bilde 2. Tjenlige mutasjoner finnes knapt
2a) 'Nesten nøytrale' mutasjoner
'Nesten nøytrale' mutasjoner har ubetydelig små effekter på genomet som et hele. Mutasjoner i nærheten av 'nesten nøytrale' nukleotide posisjoner, er automatisk emne for tilfeldig drift, som innebærer at de i all essens er immune mot naturlig seleksjon. Deres betydning for artens 'fitness' er så ubetydelig liten at den skjules av de minste fluktuasjoner eller 'støy', i biologiske systemer (Kimura, 1968, 1983, Kimura og Ohta, 1971). Disse er de mest rikelige mutasjoner, som vist i 'nesten nøytral' boksen i bildet ovenfor. Hvorfor er slike mutasjoner viktige, om de er nesten nøytrale? De har betydning fordi stedene som er utsatt for mutasjoner inneholder informasjon, og mutasjoner bidrar til å ødelegge informasjon. Kollektivt utgjør 'nesten nøytrale' mutasjoner mesteparten av informasjonsendring i genomet.
Det forholder seg på samme vis med alle, tilsynelatende ubetydelige bokstaver i denne artikkelen. Til sammen utgjør de en forhåpentlig klar melding. Om vi starter med å introdusere en laaang tekst, f.eks. i et leksikon, og vi begynner å introdusere stadig flere typografiske feil, så vil ikke enkeltfeilene ha noen særlig betydning for meningen i teksten. Men om prosessen ikke stanses, så vil meningen til slutt bli mer og mer ødelagt. Som en alternativ parallell, kan en ta rust på biler. Rust-prosessen skje atom for atom. Hvert jern-atom som oksideres synes tilsynelatende ubetydelig, men summert opp blir det en total rustskade som virker ødeleggende for bilens kjøreegenskaper og sikkerhet. En framtredende populasjons-genetiker (Kondrashov, 1995) skrev en artikkel som het: 'Hvorfor har vi ikke dødd 100 ganger?' Problemet med ikke-selekterbarhet av 'nesten nøytrale' mutasjoner er meget reelt.
Mennesker er utsatt for høyt nivå av 'støy' på genom-nivå. Vi har et stort genom, høye mutasjonsrater, høyt nivå av gen-interaksjon, og vi har meget alvorlige begrenseninger på seleksjon. Dette er hvorfor forholdet av 'nesten nøytrale' mutasjoner er meget høyt i mennesket, og frekvensen av slike mutasjoner som i realiteten er usynlige, helt skulle kontrolleres av 'tilfeldig drift'. Alle nukleotide posisjoner vil mutere fritt, og all informasjon kodet av dem vil avta på sikt.
2b. Seleksjonsterskel for 'for-mange' små mutasjoner.
 Mindre mutasjoner har liten, men distinkt effekt på reproduserende potensial. Disse er mutasjonene umiddelbart til venstre for 'nesten nøytral' boksen i figuren over. Effekten av slike mutasjoner er delvis additiv, så naturlig seleksjon kan selektere for flere slike mutasjoner samtidig. I og for seg er måten naturlige seleksjon virker på meget elegant, og virker designet for å stabilisere livet. Ellers ville det ha forfalt for lenge siden. Men problemet er at seleksjon for mindre mutasjoner har betydelige begrensninger. Eliminasjon av mindre mutasjoner blir alvorlig ødelagt av biologisk 'støy', Naturlig seleksjon kan ha problemer med å 'se' mindre mutasjoner, i og med forskjellen de utgjør på reproduksjonsevnen er meget små, mens effekten av andre faktorer kan være meget store. Det kan nesten sammenlignes med å se små bølgeformer, produsert av småstein kastet i et stormfullt hav.
Mindre mutasjoner har liten, men distinkt effekt på reproduserende potensial. Disse er mutasjonene umiddelbart til venstre for 'nesten nøytral' boksen i figuren over. Effekten av slike mutasjoner er delvis additiv, så naturlig seleksjon kan selektere for flere slike mutasjoner samtidig. I og for seg er måten naturlige seleksjon virker på meget elegant, og virker designet for å stabilisere livet. Ellers ville det ha forfalt for lenge siden. Men problemet er at seleksjon for mindre mutasjoner har betydelige begrensninger. Eliminasjon av mindre mutasjoner blir alvorlig ødelagt av biologisk 'støy', Naturlig seleksjon kan ha problemer med å 'se' mindre mutasjoner, i og med forskjellen de utgjør på reproduksjonsevnen er meget små, mens effekten av andre faktorer kan være meget store. Det kan nesten sammenlignes med å se små bølgeformer, produsert av småstein kastet i et stormfullt hav.
Bilde 3. Begrenset effekt av seleksjon
F.eks. vil seleksjon for dødelige eller nesten dødelige mutasjoner ha prioritet over de mindre effektene til mindre mutasjoner. Faktumet at de fleste mutasjoner er recessive, skjuler dramatisk deres negative effekt på fitness og hindrer seleksjon mot dem. Likedan vil all interaksjon mellom gener  (epistasis) forstyre selektiv eliminasjon av mindre mutasjoner. I mindre populasjoner, vil tilfeldigheten ved seksuell rekombinasjon rutinemessig overstyre seleksjon. Disse fenomen forårsaker det fundamentale fenomen genetisk drift. Det er velkjent at genetisk drift kan overstyre seleksjon, unntatt mot de mest alvorlige mutasjoner i små populasjoner. Alle eksterne effekter på reproduksjon vil innvirke på effektiv seleksjon. Utbredelsen av en enkelt mutasjon i en populasjon, vil tilfeldig drifte og tendere mot å bli immun mot seleksjon der netto effekten av alle andre faktorer kombinert har en større effekt på reproduktiv mulighet enn nukleotiden selv har.
(epistasis) forstyre selektiv eliminasjon av mindre mutasjoner. I mindre populasjoner, vil tilfeldigheten ved seksuell rekombinasjon rutinemessig overstyre seleksjon. Disse fenomen forårsaker det fundamentale fenomen genetisk drift. Det er velkjent at genetisk drift kan overstyre seleksjon, unntatt mot de mest alvorlige mutasjoner i små populasjoner. Alle eksterne effekter på reproduksjon vil innvirke på effektiv seleksjon. Utbredelsen av en enkelt mutasjon i en populasjon, vil tilfeldig drifte og tendere mot å bli immun mot seleksjon der netto effekten av alle andre faktorer kombinert har en større effekt på reproduktiv mulighet enn nukleotiden selv har.
Bilde 4: Kilder til fenotype variasjon. fra Kimura s.30-31
For å bruke et bilde, så kan seleksjon for meget små genetiske effekter være lik å forsøkte å høre hvisking i et stille rom. Myk hvisking, komplekse beskjeder og høy bakgrunnsstøy vil alle bidra til tap av betydning i hviskingen. Seleksjon mot mindre mutasjoner virker best når effekten den har på fitness, ennå er tydelig og bakgrunnsstøyen er minimal. Til tross for disse viktige begrensingene virker seleksjon likevel, ellers hadde vi ikke vært her. I det antall mindre mutasjoner øker, blir den individuelle effekten til hver av dem mindre og mindre betydelig (Kimura, 1983). Hver gang en ny egenskap som det må selekteres for legges til, så må maksimum seleksjonspress for hver egenskap avta. I det antall egenskaper/trekk som seleksjon rettes mot, synker effekten av seleksjon mot hvert trekk. Det er øvre rammer for hvor mange egenskaper  det samtidig kan selekteres mot. Forfatteren (J.C. Sanford) har beregnet det til å være tilnærmet 700. Kimura, som ikke viste sine beregninger, kom til at kun 138 steder kan gjennomgå seleksjon samtidig, selv for en populasjon med intenst seleksjonspress (50% eliminasjon) og meget svak selektiv eliminasjon pr. trekk (s=0,01). Ved forsøk på å selektere samtidig mot flere hundre mutasjoner, synes det naturlig om det fører til opphør av selektiv framgang.
det samtidig kan selekteres mot. Forfatteren (J.C. Sanford) har beregnet det til å være tilnærmet 700. Kimura, som ikke viste sine beregninger, kom til at kun 138 steder kan gjennomgå seleksjon samtidig, selv for en populasjon med intenst seleksjonspress (50% eliminasjon) og meget svak selektiv eliminasjon pr. trekk (s=0,01). Ved forsøk på å selektere samtidig mot flere hundre mutasjoner, synes det naturlig om det fører til opphør av selektiv framgang.
Bilde 5. Seleksjon når raskt terskel, selv ved høy fruktbarhet.
I det store bildet, i kollektive populasjoner, trengs det å selekteres mot milliarder av mutasjoner. I det antall punkter utsatt for seleksjon øker, så vil den resulterende genetiske endringen på det beste bli trukket ut i tid, og kreve hundretusener av generasjoner for å påvirke dem merkbart. Både Haldane og Kimura er enige om at det ikke samtidig er mulig simultant å påvirke et stort antall trekk, grunnet seleksjonskostnadene. Denne enkle realiteten gjør i virkeligheten genomisk seleksjon umulig.
3. Reproduktiv eliminasjon
Det foreligger også en logistikk-begrunnelse for hvorfor ikke det ikke foregår reproduktiv eliminasjon av et stort antall samtidige mutasjoner. Det kan kalles seleksjon-interferens. Problemet er ikke tilstrekkelig utredet, men er addressert av som en interferens-faktor i forbindelse med seleksjon (Haldane 1957; Lunch, Conery og Burger, 1995; Kondrashow, 1995) I forbindelse med simultan seleksjon for ti-tusener eller millioner av ulike mutasjoner i genomet, blir problemet med seleksjon-interferens overveldende.
Seleksjon-interferens oppstår når seleksjon for ett trekk henger sammen med og påvirker med seleksjon for et annet trekk. F.eks. vil et ønskelig trekk erfaringsmessig henge sammen med et ikke-ønskelig trekk i samme individ. Å selektere mot det ikke-ønskelige trekket innebærer at du automatisk må selektere mot det assosierte ønskelige trekket, i og med at seleksjon forekommer på individ-nivå. Denne forbindelsen kan være veldig nær, f.eks. to gener side om side på samme kromosom; eller være løsere to gener et eller annet sted innen et individ. Selv om to trekk bare løselig er sammenknyttet, er de likevel bundet sammen for den generasjonen. Enhver mutant vil alltid temporært være knyttet til tusener andre mutanter i hvert individ i enhver generasjon. Dermed kan seleksjon aldri isolert være knyttet til én spesifikk mutasjon.
Bilde 6. Eks. på interferens-generelt
For å illustrere dette, la oss ta et eks. mellom to individer i en populasjon. Fordi genene er knyttet til samme populasjon, vil de to individene ha omlag like mange mutasjoner og disse to sett av mutasjoner vil ha omtrent samme skadelige effekt. Vi kan f.eks. finne at hvert individ har ca. 10.000 dårlige genetiske enheter (mutasjoner), og i forhold til den andre 10.000 gode (ikke-muterte) enheter. I gjennomsnitt vil forskjellen mellom dem være liten, og bero på forskjeller i noen få nukleotider som utøver en hoved-effekt. På grunn av høy grad av total-likhet i genetisk fitness, vil reproduktiv suksess bero mer på tilfeldige endringer og 'støy-faktorer' enn på virkelig genetisk fitness. Men selv om det 'beste' individet velges ut for videreføring, så vil nærmest ingen selektiv framgang være oppnådd. Vi vil ha selektert bort 10.000 mutasjoner i det ene individet, men samtidig ha fått 10.000 ekstra mutasjoner i det andre. Eneste nettoforskjell vil gjelde for de få hoved-genene som utgjorde den faktiske forskjellen. Forskjellene vil utjamne hverandre og bli ikke-selekterbare. Selv om vi kunne ha kontroll over individer på genom-nivået, ville vi likevel feile i effektivt å forhindre videreformidling av den store masse av skadelige mutasjoner. Så vidt forfatteren (Dr. J.C. Sanford) kjenner til, er ikke dette problemet skikkelig adressert, selv om det ofte henvises til av populasjons-genetikere.
4. Seleksjons-interferens grunnet fysiske linker
Den mest tydelige og ekstreme form for seleksjon-interferens er når det er tette fysiske forbindelser mellom tjenlige og skadelige mutasjoner. Dette resulterer i et uløselig problem, referert til som 'Mullers sperreverk'. En forutsetning for at naturlig seleksjon skal kunne virke, er evnen til å separere gode og skadelige mutasjoner. Dette blir uten virkning, der de samme nevnte mutasjoner fysisk er knyttet sammen. Essensielt foreligger hele genomet i store sammenknyttede genopphopninger (clustre). (Tishkoff of Verelli, 2003) Dette problemet refererer til nær sagt  alle byggesteiner i genomet. Vi har tidligere sett at mutasjoner i all hovedsak er skadelige. De få tjenlige mutasjonene syntes imidlertid å gi en strime av håp. Men disse tjenlige mutasjoner ville i hovedsak være 'nær nøytrale', og dermed ikke-selekterbare i flg. Kimura. Dessuten ville fysiske linker mellom tjenlige og ikke-tjenlige mutasjoner, i realiteten fjerne positive mutasjoner fra vår graf for høyere genomer.
alle byggesteiner i genomet. Vi har tidligere sett at mutasjoner i all hovedsak er skadelige. De få tjenlige mutasjonene syntes imidlertid å gi en strime av håp. Men disse tjenlige mutasjoner ville i hovedsak være 'nær nøytrale', og dermed ikke-selekterbare i flg. Kimura. Dessuten ville fysiske linker mellom tjenlige og ikke-tjenlige mutasjoner, i realiteten fjerne positive mutasjoner fra vår graf for høyere genomer.
Bilde 7. Skift mot neg. mutasjonseffekt ved fysiske-linker
Fordi slike clustre aldri løses opp, så vil netto-effekten av hvert cluster arves som de besto av en enkel mutasjon. Om bare én mutasjon blant en million er tjenlig, så vil et linket par av mutasjoner hvor begge har tjenlig effekt, være så ekstremt sjeldent at det i praksis kan overses (1:1 billion). Som tiden går, vil antall mutasjoner i hver linket gruppe øke, slik at netto-tap av informasjon for hver linket gruppe vil øke, og linkede grupper med netto mutasjons-gevinst, vil forsvinne med absolutt sikkerhet. Ut fra dette kan vi si at mutasjonsoppsamlinger (clustre), f.eks. i et genom alltid er 100% skadelige. Basert på logikken ovenfor, kan vi anslå med meget høy grad av sikkerhet at disse byggestenene for evolusjon, progressivt blir av dårligere kvalitet. Basert på tallrike uavhengige slutningsrekker, er vi tvunget til å konkludere med at problemet med menneskelig genom-degenerering er reelt. utslokking av menneskelig genom synes å være like sikkert som stjernenes utslokking, organismers død og universets opphør en gang. Langvarig genom-stabilitet er bare mulig om det er en ukjent, men fundamental kraft som motvirker genomisk entropi.
Tillegg: I 2006 kom det en artikkel hvor det utfra Mullers 'sperreverk', ble argumentert for en dødelig effekt for menneskelig rase innenfor evolusjonær tidsskala. Det skjedde utfra en enkelt sammenføynings-enhet, mitokondrie-kromosomet (Loewe, 2006). Men dette er bare én av ca. 200.000 linkede grupper i menneskelig genom.
Oversatt og gjort om til .htm-format ved Asbjørn E. Lund