Problemer knyttet til seleksjon
-Tre spesifikke seleksjonsproblemer
( Fra: 'Genetic Entropy & the Mystery of the Genome; Dr. J.C.Sanford; FMS Publications; Ch. 4-slutten)
 Vi skal se på en enkel problemstilling i starten: Anta at vi har en punktmutasjon som har bredt seg til 50% av befolkningen. Hvilken type seleksjon kreves for å eliminere denne mutasjonen, og hva er kritiske faktorer i så måte? For enkelhets skyld vil vi anta at mutasjonen er dominant. Det faktum at nesten samtlige mutasjoner er recessive, gjør det mye vanskeligere for seleksjon å luke dem vekk. I en matematisk tilnærming, evt. for ‘Hitler-aspiranter’ er problemet enkelt løst: Vi kunne eliminere samtlige mutasjoner i én generasjon, om vi kunne miste 50% av den reproduserende populasjonen. ‘Så hva er problemene?’
Vi skal se på en enkel problemstilling i starten: Anta at vi har en punktmutasjon som har bredt seg til 50% av befolkningen. Hvilken type seleksjon kreves for å eliminere denne mutasjonen, og hva er kritiske faktorer i så måte? For enkelhets skyld vil vi anta at mutasjonen er dominant. Det faktum at nesten samtlige mutasjoner er recessive, gjør det mye vanskeligere for seleksjon å luke dem vekk. I en matematisk tilnærming, evt. for ‘Hitler-aspiranter’ er problemet enkelt løst: Vi kunne eliminere samtlige mutasjoner i én generasjon, om vi kunne miste 50% av den reproduserende populasjonen. ‘Så hva er problemene?’
Bilde 1. Teoretisk grunnlaget for naturlig utvalg. Se mer her.
Seleksjonskostnader
Seleksjon har i våre dager lett for å bli et mantra, som kan løse alle problemer knyttet til evolusjon. Få tenker over at dette har en kostnad i en populasjon. At seleksjon faktisk innebærer å velge bort noe, er baksiden av medaljen som skrivebords-teoretikere lett kan overse. Poenget med denne gjennomgangen er å vise at det er klare grenser for hvor intens seleksjonen kan være, i en menneskelig populasjon som (forhåpentlig) skal overleve på sikt. Situasjonen er totalt annerledes i forhold til en populasjon med høy fruktbarhet; f.eks. i en plantepopulasjon kunne en ‘forbruke’ 99% av formerende populasjon til selektive formål, på grunn av ekstrem forplantingsdyktighet (hundre folds).
Når vi skal betrakte genetiske faktorer som bestemmer reproduksjons-suksessen, så er det et poeng at enkelte viktige genetiske trekk ikke bringes videre til avkommet. F.eks. virker mange gener bra i visse kombinasjoner, men er ikke ønskelige i og for seg selv. Seleksjon på slike genkombinasjoner, er i virkeligheten falsk seleksjon fordi det ikke nødvendigvis medfører noe godt for avkommet. En vet ikke hvilke kombinasjoner som da blir ‘gode’, i kombinasjon med andre gener etc. Slik ‘falsk seleksjon’ må likevel betales for, noe som krever enda høyere reproduksjon. Og vi har enda ikke begynt å se på reell seleksjon.
Seleksjon kan anta flere former: stabiliserende seleksjon, seksuell seleksjon, progressiv seleksjon etc. Hver type seleksjon har en kostnad som er additiv og må betales for. De totale reproduktive kostnadene må aldri overstige det aktuelle reproduksjons potensialet hos arten. Bare om en art er tilstrekkelig fruktbar og det er tilstrekkelig overskudds populasjon til å dekke alle andre kostnader, er det mulig med noen form for seleksjon. Ellers vil populasjonen avta og dø ut. Det blir litt likt skjønnsomt forbruk i en husholdning: spørsmålet blir alltid om en har råd til det.
Vi skal i det følgende først betrakte kunstig seleksjon, før vi går over til naturlig:
1. Prosentdel av mennesker tilgjengelig for selektiv fjerning
Bare en begrenset andel av populasjonen (populasjons-overskuddet) kan bli valgt bort selektivt, uten at populasjonen til slutt dør ut. (Eks. nettoreproduksjonstallet 'antall levendefødte jenter, hver kvinne i alderen 15–44 år kommer til å føde under gjeldende fruktbarhets- og dødelighetsforhold'. Dette tallet var i Norge 0,86 i 2003. For å opprettholde folketallet på lengre sikt må det være 1,0)
Globalt blir det født mindre enn 3 barn for 2 voksne (foreldre). Fødselsoverskuddet er på ca. 1/3. Ut fra dette må 2/3 av barna reprodusere for å erstatte de som dør. Av den resterende 1/3 del, er det en signifikant andel som av ulike grunner ikke reproduserer. Det skyldes alt fra sykdom, krig, ulykker, personlige valg etc. som ikke har med fenotypen å gjøre. I visse deler av verden, kan det faktisk utgjøre halvparten av denne 1/3-delen. Dermed blir det igjen bare omkring 1/6 (16,7%) som faktisk er tilgjengelig for potensiell selektivt bort-velging.
Bilde 2. Tilgjengelig andel for sekeksjon (16,7%)
Fitness er betegnelse for et individs evne til å få avkom, sett i forhold til hvor mange avkom andre individer får. Fitness avhenger altså både av individets overlevelses- og formeringsevne. Jo høyere fitness et individ har, dess høyere sannsynlighet har det for å bli selektert. Fitness i forhold til fenotype-overlegenhet, er alltid det trekk naturlig seleksjon handler i forhold til. Det er et fundamentalt at dette trekket er veldig lite arvbart, i motsetning til populær tenkemåte omkring det. I følge Kimura har fitness kanskje så lav arvelighet som 0,004 (Kimura ,1983, s30-31). Det innebærer at omgivelses-faktorer er mye viktigere enn genetiske faktorer i å bestemme hvem som framstår som 'overlegen'. Det at Kimura sier at generell fitness har veldig lav arvelighet, er en forbløffende erkjennelse. Det innebærer at seleksjon for generell overlegenhet (fitness) har meget liten betydning for sammensetningen av neste generasjon.

Bilde 3 Antall levende fødte pr. 1000 kvinner i 5-års-grupper
Forfatteren (J.S. Sanford) tror at en av de mest fundamentale feil som teoretikere gjør, i det de finner opp sine ulike scenarioer er å ignorere seleksjons-kostnader. De opptrer som det alltid er en stor overskuddsbefolkning. De opptrer ofte som de er ubundet av realistiske kostnadsrammer. Dermed kan de 'betale for' et hvilket som helst omfang av simultane seleksjons-scenarier. Om en holdt seg til realiteten i stedet for fantasier, ville det innebære mye mer realistiske forventninger om hva seleksjon virkelig kan utføre.
2. Gjenkjente fordunklede (usynlige) mutasjoner
Om du ønsker å selektere i forhold til en mutant, må du være i stand til å identifisere den innen populasjonen. Men for å identifisere bærer av en typisk punktmutasjon, trenger en å screene en hel populasjon, noe som er kostbart og ikke realistisk. De fleste punktmutasjoner er som nevnt så hårfine at de i praksis er usynlige, selv om de er destruktive og fører til tap av informasjon. Når en skal vurdere millioner av mutasjoner samtidig, og deretter velge mellom to personer, vil det nødvendigvis føre til vilkårlige valg.
3. Systematisk reproduktiv eliminasjon
 Seleksjonskostnader og stile mutasjoner er store problemområder som blir enda større om en skal hindre mutante individer å formere seg. Det finnes knapt noe sted på planeten der en kan kontrollere menneskelig reproduksjon med høy presisjon. Det mest beryktede eksempel er Nazi-Tysklands forsøk under 2. verdenskrig. Heldigvis feilet det eksperimentet katastrofalt. Selv om en i familieplanlegging har gjennomført en hard linje i Kina, har en ikke eliminert mutasjoner. I de fleste tilfelle skjer menneskelig formering og reproduksjon temmelig tilfeldig, unntatt i meget sjeldne tilfelle hvor mutasjoner resulterer i meget uttalte genetiske defekter.
Seleksjonskostnader og stile mutasjoner er store problemområder som blir enda større om en skal hindre mutante individer å formere seg. Det finnes knapt noe sted på planeten der en kan kontrollere menneskelig reproduksjon med høy presisjon. Det mest beryktede eksempel er Nazi-Tysklands forsøk under 2. verdenskrig. Heldigvis feilet det eksperimentet katastrofalt. Selv om en i familieplanlegging har gjennomført en hard linje i Kina, har en ikke eliminert mutasjoner. I de fleste tilfelle skjer menneskelig formering og reproduksjon temmelig tilfeldig, unntatt i meget sjeldne tilfelle hvor mutasjoner resulterer i meget uttalte genetiske defekter.
Bilde 4. Nye (spillgale) tider gir nye seleksjonskriterier
Fra våre betraktninger må vi slutte at vi ikke er i stand til å selektere kunstig for endog én punktmutasjon innen menneske-rasen. Om vi går videre til multiple mutasjoner, eskalerer problemnivået eksponentielt. Selv om en i teorien kunne identifisere alle bærere av mutasjoner og hindre dem i å reprodusere, ville en snart løpe inn i problemer med en raskt synkende populasjons størrelse. Seleksjon er ikke så enkelt i praksis som i teorien. Selv for det enkleste seleksjons-scenario kreves følgende viktige faktorer: 1) Vedlikehold av populasjons-størrelsen. 2) Entydig identifikasjon av mutanter og 3) effektiv utelukking av mutanter fra formerende populasjon. Når en vurder alle mutasjoner samlet sett, så blir hvert eneste av disse tre kravene en umulighet.
Etter å ha sett på problemer med kunstig seleksjon, er det grunn til å spørre om naturlig seleksjon kommer populasjonen til redning? Men naturlig seleksjon møter akkurat de samme problemene. En møter de samme seleksjonskostnadene: En kan ikke selektere mot for mange mutasjoner samtidig, da vil den bli ineffektiv eller føre til redusert populasjons-størrelse. Om en samtidig skal selektere bort mengder av mutasjoner, treffer en på problemet med seleksjons-interferens (samvirke). Disse meget realistiske beskrankninger på naturlig seleksjon, begrenser hva vi realistisk kan forvente at naturlig seleksjon skal fullende.
 I et begrenset omfang vil likevel genetisk seleksjon ha sin effekt. ‘Genetisk engineering’ i forhold til planter, benytter seleksjonsteknikker for å få genetisk tillagde planter til å komme seg. Likedan har naturlig seleksjon valgt bort de verste menneskelige mutasjoner. Ellers hadde menneske-rasen degenerert for lenge siden. Men både naturlig og kunstig seleksjon har meget begrensede områder til sin disposisjon. Selv om naturlig seleksjon kan danne noen spesifikke gen-frekvenser, så kan ingen seleksjon vedlikeholde, langt mindre danne, høyere genom. For korthets skyld kan vi konkludere slik: Seleksjon kan virke noen ganger på genetisk nivå, men feiler systematisk på genomnivået.
I et begrenset omfang vil likevel genetisk seleksjon ha sin effekt. ‘Genetisk engineering’ i forhold til planter, benytter seleksjonsteknikker for å få genetisk tillagde planter til å komme seg. Likedan har naturlig seleksjon valgt bort de verste menneskelige mutasjoner. Ellers hadde menneske-rasen degenerert for lenge siden. Men både naturlig og kunstig seleksjon har meget begrensede områder til sin disposisjon. Selv om naturlig seleksjon kan danne noen spesifikke gen-frekvenser, så kan ingen seleksjon vedlikeholde, langt mindre danne, høyere genom. For korthets skyld kan vi konkludere slik: Seleksjon kan virke noen ganger på genetisk nivå, men feiler systematisk på genomnivået.
Nærmere om kilder til fenotype variasjoner
( Fra: 'Genetic Entropy & the Mystery of the Genome; Dr. J.C.Sanford; FMS Publications; s100-102)
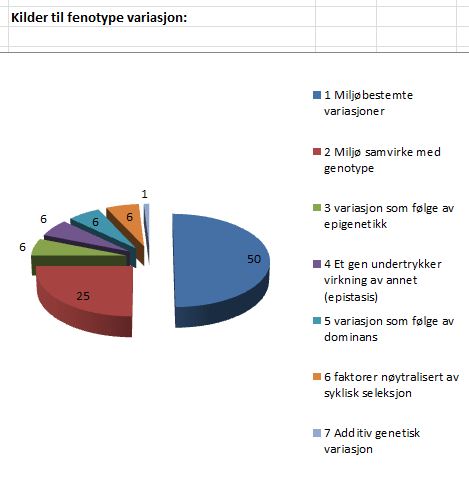
Bilde 5. Kilder til fenotype variasjon. fra Kimura s.30-31
Variasjon mellom individer kommer fra tallrike genetiske og ikke-genetiske faktorer. Innen den genetiske komponenten, er det også flere komponenter. Bare en genetisk komponent er selekterbar. Men denne komponenten er helt ‘overdøvet’ av andre kilder til fenotype variasjon. Hovedkilden (1) til fenotype variasjoner er miljøbestemte variasjoner (50%). Denne variasjonen er ikke (kjent som) arvbar, og samvirker med seleksjon. En annen hovedkilde (2) er samvirke mellom miljøfaktorer og genotype (25%) . Innen den genetiske variasjons-komponenten er (3) variasjon som skyldes epigenetikk (ca.6%) {-som kan være arvelige -oversetters kommentar}. (4) Epistasis: samhandling mellom to eller flere gener, for å kontrollere én bestemt fenotype, spesielt ved at ett gen stopper effekten av det andre. (ca.6%). Så har vi (5) variasjon via dominans (ca.6%). Ingen av disse faktorene er arvbare, og de samvirker med seleksjon. Så er det andre genetiske faktorer, som kunne vært selekterbare, men av ulike grunner er nøytralisert. F.eks. av (6) homeostatiske prosesser eller slikt som syklisk seleksjon (ca.6%). Dette etterlater (7) arvelige additiv genetisk variasjon, som en relativt ubetydelig komponent for fenotype-variasjon (ca.1%)

Prosentdel av mennesker tilgjengelig for selektiv fitness-utvalg
Selv om det er en teoretisk andel på 16,7 % som er tilgjengelig for seleksjon, gitt at populasjonen skal bestå (Bilde 2), så er det statistisk bare 0,4%(=0,004) av en slik seleksjon, som vil resultere i arvelige effekter hos påfølgende generasjoner. Dette innebærer en arvelig effekt hos mindre enn 0,1% Prosentvis: 16,7%*0,004= 0,0668%, altså mindre enn 0,1%.
Det innebærer at mindre enn 1 person pr. 1000 kan selekteres bort for effektiv, selektiv reproduktiv eliminasjon. Selv om en ‘Hitler-type’ skulle fjerne så mange ‘mindreverdige individer’ som mulig, så ville det resultere i en ikke-signifikant selektiv prosess for noe så generelt som egnethet (fitness). Det er en konklusjon av lav menneskelig fruktbarhet og den ekstremt dårlige arveligheten (0,4%) av en slik egenskap.
Bilde 6. Tilgjengelig andel for fitness-utvalg
Tekst og diagrammer tilrettelagt på norsk ved Asbjørn E. Lund