Darwinisme og felles avstamning
(oppsamlet fra 'info-krever-intelligens.info/blogg' om emnet. Opprinnelig hentet fra bøkene : 'Darwin 200 år' og 'Guds bøddel')
Evolusjonsmyten
 Det fins mange myter innen darwinismen, men den viktigste er avstamningsmyten. I følge denne myten vil absolutt alle organismer som har levd på jorden til alle tider, henge sammen i et felles naturlig og kjødelig avstamningsbånd. Darwinister innser at det er vanskelig å tro på dette felles kjødelige båndet. Derfor sier de at denne kjeden ikke eksisterer fordi de ulike leddene i den ikke lever samtidig. Men det gir like lite mening som å si at jeg ikke lenger er i slekt med mine søsken, fra den dag foreldrene våre var døde. Evolusjonsmyten bygger på at jeg og alle andre levende vesener har en tipp-tipp-tipp-...-tipp stamfar i urcellen i 'ursuppen'.
Det fins mange myter innen darwinismen, men den viktigste er avstamningsmyten. I følge denne myten vil absolutt alle organismer som har levd på jorden til alle tider, henge sammen i et felles naturlig og kjødelig avstamningsbånd. Darwinister innser at det er vanskelig å tro på dette felles kjødelige båndet. Derfor sier de at denne kjeden ikke eksisterer fordi de ulike leddene i den ikke lever samtidig. Men det gir like lite mening som å si at jeg ikke lenger er i slekt med mine søsken, fra den dag foreldrene våre var døde. Evolusjonsmyten bygger på at jeg og alle andre levende vesener har en tipp-tipp-tipp-...-tipp stamfar i urcellen i 'ursuppen'.
Darwinismens årsaker kommer nedenfra
 Noen av oss lærte som barn at 'alt det gode kommer ovenfra'. Grunntanken i darwinismen er motsatt: 'Alle årsaker kommer nedenfra'. I darwinistisk tankegang er ikke livet noe som skjenkes oss ovenfra, tvert i mot kommer det fra pølen der nede. Livet kommer snikende fra en uidentifiserbar amøbe, nede i 'ursuppa'. Denne amøben har dratt seg opp etter håret den ikke har, og er blitt mer og mer avansert på sin vei til å bli menneske. Det må være et filosofisk og logisk problem for darwinismen at alle forklaringer er fattigere enn fenomenene de skal forklare. Ved stadig reduksjon blir uvegerlig noe borte underveis. Det at liv spontant skal ha blitt til av ikke-liv er et slikt eksempel.
Noen av oss lærte som barn at 'alt det gode kommer ovenfra'. Grunntanken i darwinismen er motsatt: 'Alle årsaker kommer nedenfra'. I darwinistisk tankegang er ikke livet noe som skjenkes oss ovenfra, tvert i mot kommer det fra pølen der nede. Livet kommer snikende fra en uidentifiserbar amøbe, nede i 'ursuppa'. Denne amøben har dratt seg opp etter håret den ikke har, og er blitt mer og mer avansert på sin vei til å bli menneske. Det må være et filosofisk og logisk problem for darwinismen at alle forklaringer er fattigere enn fenomenene de skal forklare. Ved stadig reduksjon blir uvegerlig noe borte underveis. Det at liv spontant skal ha blitt til av ikke-liv er et slikt eksempel.
Avstamningshypotesen er en tanke om avstamning nedenfra, fra primitivt til høyerestående liv. Mutasjoner er en 'årsak' som kommer nedenfra, fordi den forstyrrer alle bestående strukturer. Naturlig utvalg er en 'årsak' som kommer nedenfra, fordi den prøver forklare ting bare ved å velge bort egenskaper fra bestående mangfold. Altså er darwinisters hovedforklaring ikke forklaring i det hele tatt.
 For kunst og musikk kan det medvirke til at en tror at verket er rikere enn kunstnerens sinn, eller rikere enn fargesammensetningene til rådighet på paletten. En kan innbille seg at komposisjonen holder et høyere musikalsk nivå enn komponisten. En tror maleriet blir vakrere enn forestillingen kunstneren hadde i sitt hode, da han laget det. Denne tankegangen har et filosofisk forklaringspoblem: Noe framstilles uten fullverdig årsak, -hvor kommer 'resten' fra? Men dette materialistiske prosjektet er ikke så viselig at det lar seg gjennomføre konsekvent..
For kunst og musikk kan det medvirke til at en tror at verket er rikere enn kunstnerens sinn, eller rikere enn fargesammensetningene til rådighet på paletten. En kan innbille seg at komposisjonen holder et høyere musikalsk nivå enn komponisten. En tror maleriet blir vakrere enn forestillingen kunstneren hadde i sitt hode, da han laget det. Denne tankegangen har et filosofisk forklaringspoblem: Noe framstilles uten fullverdig årsak, -hvor kommer 'resten' fra? Men dette materialistiske prosjektet er ikke så viselig at det lar seg gjennomføre konsekvent..
Det naturlige utvalg som årsak
 Det er lov å prøve seg. Men det å tro at en forstår noe er ikke ensbetydende med at en faktisk gjør det. Biologiprofessorene har rett i at det er mulig å forstå det naturlige utvalg. Det naturlige utvalg har bare et trikk til rådighet: ' Å velge bort ting.' Det naturlige utvalgs repertoar kan sammenlignes med trubaduren Paul Simons 'One Trick Pony'. Denne ponnien hadde kun ett trick til rådighet der den reiste rundt og viste seg fram. Dermed nådde den ikke den store populariteten, men det naturlige utvalg har blitt den store vinneren! Om det ikke hjelper oss å forstå naturen, så har det i hvert fall hjulpet darwinismen å bli vinneren så langt. Men det kan snu! Tricket til ponnien var sikkert mer avansert enn det naturlige utvalg kan varte opp med.
Det er lov å prøve seg. Men det å tro at en forstår noe er ikke ensbetydende med at en faktisk gjør det. Biologiprofessorene har rett i at det er mulig å forstå det naturlige utvalg. Det naturlige utvalg har bare et trikk til rådighet: ' Å velge bort ting.' Det naturlige utvalgs repertoar kan sammenlignes med trubaduren Paul Simons 'One Trick Pony'. Denne ponnien hadde kun ett trick til rådighet der den reiste rundt og viste seg fram. Dermed nådde den ikke den store populariteten, men det naturlige utvalg har blitt den store vinneren! Om det ikke hjelper oss å forstå naturen, så har det i hvert fall hjulpet darwinismen å bli vinneren så langt. Men det kan snu! Tricket til ponnien var sikkert mer avansert enn det naturlige utvalg kan varte opp med.
Darwinister har laget sin egen seleksjonsliturgi  der følgende trosbekjennelse framføres: 'Denne egenskapen (seleksjon) utviklet seg fordi det var gunstig for overlevelse. I NRKs naturprogram på TV, ble ordet overlevelse på brukt på en spesiell måte: 'overlevelse i form av seier i konkurransen om økologiske nisjer. I følge darwinismen må alltid seleksjonen være årsak til spesielle egenskaper hos en art. Men la oss se litt grundigere på dette. Det kan faktisk finnes andre og flere muligheter: i) En annen og dypere årsak kan ligge bak det naturlige utvalg. ii) Det naturlige utvalg kan være en virkning i stedet for en årsak. iii) Det naturlige utvalg er verken en årsak eller en virkning.
der følgende trosbekjennelse framføres: 'Denne egenskapen (seleksjon) utviklet seg fordi det var gunstig for overlevelse. I NRKs naturprogram på TV, ble ordet overlevelse på brukt på en spesiell måte: 'overlevelse i form av seier i konkurransen om økologiske nisjer. I følge darwinismen må alltid seleksjonen være årsak til spesielle egenskaper hos en art. Men la oss se litt grundigere på dette. Det kan faktisk finnes andre og flere muligheter: i) En annen og dypere årsak kan ligge bak det naturlige utvalg. ii) Det naturlige utvalg kan være en virkning i stedet for en årsak. iii) Det naturlige utvalg er verken en årsak eller en virkning.
Vi vet at at recessive gener ofte er den dypere og egentlige årsak bak fenomener, som kan se ut som resultat av det naturlige utvalg. Det naturlige utvalg er bare en metode for å lokke fram egenskaper som ligger godt gjemt i artens arvestoff. Det naturlige utvalg er heller ingen årsak til å overvinne i konkurransen om økologiske nisjer. Det er heller motsatt. Det er konkurransen om begrensede goder, som er den bakenforliggende årsaken. Og det naturlige utvalg er virkningen av denne årsaken. For å fylle økologiske nisjer er det et annet prinsipp som gjelder: 'førstemann til mølla, er den som får male først'. Det er først når konkurransen tetner seg til at det naturlige utvalg trår i kraft. Det naturlige utvalg er heller ikke årsak til livets opprinnelse. Der måtte det noe kvalitativt nytt til.
Destruktive krefter som ikke kan skape
 Så kan en alltids undres hvorfor darwinismen har så lite konkret å bidra med, når det er kriser i naturen (f.eks. bi-døden). Etter vår mening er det fordi darwinismen er en tankeflukt. En akrobatisk øvelse i å tenke i strid med virkeligheten. Darwinismens så såkalte mekanismer snur tingene fullstendig på hodet når det gjelder byggende kontra destruktive krefter i tilværelsen:
Så kan en alltids undres hvorfor darwinismen har så lite konkret å bidra med, når det er kriser i naturen (f.eks. bi-døden). Etter vår mening er det fordi darwinismen er en tankeflukt. En akrobatisk øvelse i å tenke i strid med virkeligheten. Darwinismens så såkalte mekanismer snur tingene fullstendig på hodet når det gjelder byggende kontra destruktive krefter i tilværelsen:
a) Naturlig utvalg er 'borgerkrig'. En nedbrytende kraft som er i stand til å ensrette og gradvis redusere det rike mangfold som naturen er utrustet med. Så hjelper det lite om det finnes andre krefter som kan gjøre dette enda raskere.
b) Tilfeldige mutasjoner er støy. En forstyrrende kraft som gradvis bryter ned den finstemte informasjonen, harmonien og balansen som er nedlagt i skaperverket.
 Egentlig er det ganske hårreisende at det er mulig å gjøre disse negative kreftene positivt ansvarlig for alt liv som utfolder seg på planeten. I stedet for å reise bust mot denne pseudo-åndelige, kvasi-intellektuelle tankegangen, bøyer mange hodet uten å mukke under det destruktive åket. Teologer av god vilje hevder på ramme alvor at kirkens skapelses-formidling må være et lydrike under darwinismens mytologi. Og det er en mytologi på kollisjonskurs med realitetene. La oss se mer på hvordan darwinismens to 'mekanismer' blir tillagt egenskaper stikk motsatt av det de har:
Egentlig er det ganske hårreisende at det er mulig å gjøre disse negative kreftene positivt ansvarlig for alt liv som utfolder seg på planeten. I stedet for å reise bust mot denne pseudo-åndelige, kvasi-intellektuelle tankegangen, bøyer mange hodet uten å mukke under det destruktive åket. Teologer av god vilje hevder på ramme alvor at kirkens skapelses-formidling må være et lydrike under darwinismens mytologi. Og det er en mytologi på kollisjonskurs med realitetene. La oss se mer på hvordan darwinismens to 'mekanismer' blir tillagt egenskaper stikk motsatt av det de har:
1) Tilfeldige mutasjoner får æren for oppbygningen av biologiens orden og genetiske informasjon. I virkeligheten virker mutasjonene ensidig og irreversibelt nedbrytende på biologisk orden og genetisk informasjon.
2) Seleksjonen tildeles ansvar for livets fantastiske mangfold. I virkeligheten foretar det naturlige utvalg en ensidig og irreversibel reduksjon av naturens mangfold.
Det er en merkelig retorikk som gjør seg gjeldende omkring naturlig utvalg. Om en hevder at naturlig utvalg bidrar til å redusere mangfold, kan noen bli røde av sinne. Da kan det være godt å minnes at det er menneskeheten som er den eneste art som kan rødme. Når naturlig seleksjon påstås å bringe egenskaper inn i en populasjon, skyldes det som nevnt recessive egenskaper i populasjonen. Når egenskaper i populasjonen kommer for langt på grunn av seleksjon, kan det bare gå én vei: ut av populasjonen. Endelig tar det naturlige utvalg fokus vekk fra overlevelse og flytter det over til en fiktiv perfeksjonisme, mens selve livet tas for gitt. Det reduserer til noe en 'tar for gitt'.
Digital og analog
Sidens også biologien bærer stadig mer preg av å bli en digital vitenskap, tar vi med litt om ulikheten mellom analog og digital. Ordet digital kommer fra latin 'digitus', som betyr finger. Det stammer altså fra å telle på fingrene, og grunnbetydningen av digital blir dermed 'heltallig'. En digital graf i et diagram kan betegnes som følgende: trinnvis, disket,  diskontinuerlig. Et digitalt diagram kalles gjerne for et histogram, når det framstilles i form av søyler.
diskontinuerlig. Et digitalt diagram kalles gjerne for et histogram, når det framstilles i form av søyler.
Ordet analog betyr i dagligtale 'tilsvarende' eller 'lignende'. Det blir for upresist for vitenskapelig bruk. I teknologisk sammenheng brukes analog som motsetning til digital. Den brukes da i betydningen kontinuerlig. En analog størrelse kan endre seg jevnt, uten store sprang. En analog kurve i et diagram betegnes ofte som glatt, jevn eller kontinuerlig. En kan oppnå en kontinuerlig kurve ved å tilpasse den til midtpunktet i søylenes toppflate i et histogram. Ved store observasjonssett (N>1000) kan en regne med 'de store talls lov', som vil jevne ut observasjonsverdiene.
Tiden som faktor
 Det er ingen grunn til å tro at tiden skulle følge 'de store talls lov', som gjør at en kan regne med 'kontinuerlige endringer'. En gjetter på at tiden skal sørge for en glatt overgang fra art til art basert på dogmatisk tro, ikke på empiri. Dette er en fullstendig uoppgivelig tro for en darwinist, og har blitt en utbredt oppfating i Vesten. Riktignok kan det være dem som hevder dette av praktiske eller taktiske grunner: En orker rett og slett ikke ta opp kampen mot noe som virker så inngrodd som dette. Vi gjør likevel et forsøk på å klargjøre vår posisjon her! Men det er også mye annet å gripe fatt i når det gjelder darwinismen..
Det er ingen grunn til å tro at tiden skulle følge 'de store talls lov', som gjør at en kan regne med 'kontinuerlige endringer'. En gjetter på at tiden skal sørge for en glatt overgang fra art til art basert på dogmatisk tro, ikke på empiri. Dette er en fullstendig uoppgivelig tro for en darwinist, og har blitt en utbredt oppfating i Vesten. Riktignok kan det være dem som hevder dette av praktiske eller taktiske grunner: En orker rett og slett ikke ta opp kampen mot noe som virker så inngrodd som dette. Vi gjør likevel et forsøk på å klargjøre vår posisjon her! Men det er også mye annet å gripe fatt i når det gjelder darwinismen..
Darwinistisk tidsjuks
 Det foreligger en vandrehistorie blant darwinister om at mengder med fossilfunn er blitt presist forutsagt av darwinister selv. Dette kan knapt være lenger unna realiteten. De har aldri hatt fjerneste anelse om hvilke nye fossiler som kan komme rundt neste sving. Det som har skjedd derimot, er at mengder med fossilfunn er blitt tolket i etterpåklokskap, med eller uten rette, av darwinister. En sier en visste at akkurat disse funnene ville komme. Men først etter å ha funnet dem. Den ledende engelske paleontologen Colin Patterson (1933-1998) avslører hvorfor det står slik til. Han innrømte at fossile avstamningsrekker går utelukkende på fossilenes forhold til nålevende arter. Stamtavlene som settes opp, er bestemt av hvert fossils likhet med nålevende arter. Stamtavlene handler ikke om fossilenes forhold til andre fossiler. Og dette er et 'slektskap' i form av synlig anatomisk likhet (homologi). Det er ikke et slektskap i form av felles avstamning, for en vet uansett svært lite om hvem som evt. måtte stamme fra hvem i fortiden
Det foreligger en vandrehistorie blant darwinister om at mengder med fossilfunn er blitt presist forutsagt av darwinister selv. Dette kan knapt være lenger unna realiteten. De har aldri hatt fjerneste anelse om hvilke nye fossiler som kan komme rundt neste sving. Det som har skjedd derimot, er at mengder med fossilfunn er blitt tolket i etterpåklokskap, med eller uten rette, av darwinister. En sier en visste at akkurat disse funnene ville komme. Men først etter å ha funnet dem. Den ledende engelske paleontologen Colin Patterson (1933-1998) avslører hvorfor det står slik til. Han innrømte at fossile avstamningsrekker går utelukkende på fossilenes forhold til nålevende arter. Stamtavlene som settes opp, er bestemt av hvert fossils likhet med nålevende arter. Stamtavlene handler ikke om fossilenes forhold til andre fossiler. Og dette er et 'slektskap' i form av synlig anatomisk likhet (homologi). Det er ikke et slektskap i form av felles avstamning, for en vet uansett svært lite om hvem som evt. måtte stamme fra hvem i fortiden
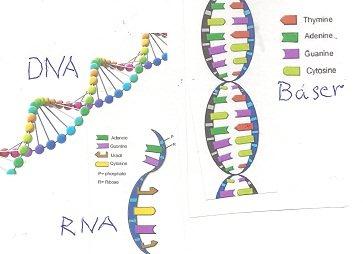
Misforståelser om DNA:
Noen mener at naturlig avstamning er naturlig fordi alle levende vesener har genetikk basert på DNA.
 a) DNA er ingen genetisk kode. DNA er bare et kodingsmedium, på samme måte som trykksverten i bøker er det.
a) DNA er ingen genetisk kode. DNA er bare et kodingsmedium, på samme måte som trykksverten i bøker er det.
b) Bokstavene i en bok svarer til de fire ulike formene for trappetrinn, som holder DNA-dobbeltspiralen sammen (AT,TA, CG og GC)
c) Darwin visste ingenting om DNA og hadde ikke noe grunnlag for å gjette på en ubrutt kontinuitet mellom ulike livsformer.
d) Livsformenes felles medium, DNA, antyder ganske riktig et felles opphav. Men en overordnende intelligens, eller en felles skaper, er det hakk mer logisk enn antagelsen om kontinuitet tilbake til en ikke-skapt urcelle, som bare fant på å oppstå av seg selv.
e) DNA mediet benytter digital koding, som ikke kan endres kontinuerlig fra art til art. Kontinuiteten må i så fall være der i utgangspunktet, i form av en felles hybridisering.
f) Alle fossilfunn svarer til punkter i et diagram av typen kompleksitet mot tid. Punkter i diagrammet er som hull av haglkorn i en papplate. Avstamningshypoteser svarer til å trekke linjer mellom haglene. Ingen vil vel finne på å si at alle hull etter blyhagl egentlig danner samsvarende linjer.
Om digitale algoritmer
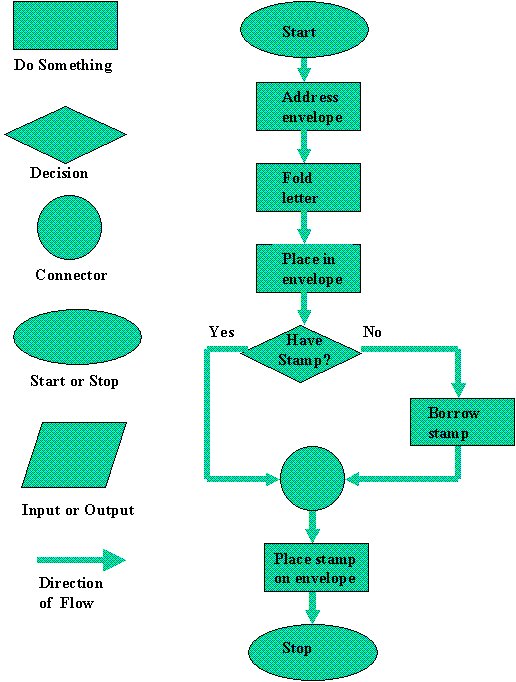 Ordet algoritme kommer fra et arabisk ord 'algawarizmiat', som i dagligspråket kan omsettes med 'oppskrift'. Definisjonen lyder: 'en algoritme er en endelig sekvens med presise instruksjoner, som skal utføres i rekkefølge.' Inkludert i slike algoritmer kan det være tester, hvor resterende instruksjoner avhenger av testen. Det kan være løkker, hvor en gjentar instruksjoner et visst antall ganger. Algoritmer kan med fordel beskrives grafisk, ved flytdiagrammer (flow-charts). De starter gjerne med en start-tilstand og ender i en slutt-tilstand. Algoritmer kan utføres på en digital datamaskin ved hjelp av programmeringsspråk. Da må algoritmen først formuleres på i programvare (formuleringsnivå) og siden utføres (eksekveres) ved hjelp av maskinvare (hardware).
Ordet algoritme kommer fra et arabisk ord 'algawarizmiat', som i dagligspråket kan omsettes med 'oppskrift'. Definisjonen lyder: 'en algoritme er en endelig sekvens med presise instruksjoner, som skal utføres i rekkefølge.' Inkludert i slike algoritmer kan det være tester, hvor resterende instruksjoner avhenger av testen. Det kan være løkker, hvor en gjentar instruksjoner et visst antall ganger. Algoritmer kan med fordel beskrives grafisk, ved flytdiagrammer (flow-charts). De starter gjerne med en start-tilstand og ender i en slutt-tilstand. Algoritmer kan utføres på en digital datamaskin ved hjelp av programmeringsspråk. Da må algoritmen først formuleres på i programvare (formuleringsnivå) og siden utføres (eksekveres) ved hjelp av maskinvare (hardware).
Når vi nå vet at livet er algoritmisk og digitalt, kommer det i strid med darwinismens påstand om en kontinuerlig avstamning fra urcelle til menneske. I livets algoritmer er det kun sprangvise, ikke gradvise (kontinuerlige) endringer, evt. er det uovertigelige barrierer. Digitale algoritmer er i ferd med å revolusjonere moderne teknologi og biologi. Men vitenskapelig sett innebærer algoritmer viktige begrensninger som skaper uoverstigelige problemer for darwinismen:
 i) Alle kjente algoritmer må antas å være designet. En har ennå ikke funnet entydige algoritmer, som er blitt til av seg selv.
ii) Alle algoritmer er hierarkiske. De inneholder minimum et formuleringsnivå (software) og et eksekveringsnivå (hardware).
i) Alle kjente algoritmer må antas å være designet. En har ennå ikke funnet entydige algoritmer, som er blitt til av seg selv.
ii) Alle algoritmer er hierarkiske. De inneholder minimum et formuleringsnivå (software) og et eksekveringsnivå (hardware).
iii) Progresjonen i en algoritme, består vanligvis av ikke-ombyttbare instruksjoner.
iv) De grunnleggende livsprosessene består oftest av entydige algoritmer. (Det finnes noen unntak med slingringsmonn. Mest i forbindelse med kjønnet formering, som må være robust for at arten skal overleve.)
v) Funksjonelle algoritmer bør oppfattes som en helhet. En biologisk algoritme må vanligvis være fullstendig før den kan iverksettes.
vi) Algoritmer må starte i en spesifisert start-tilstand med bestemte parametervalg (aldri fra et nullpunkt). En funksjonell algoritme danner oftest en ubrytelig helhet. Mange algoritmer kan ikke bygges ut trinn for trinn.
 vii) Mutasjoner forutsetter at ferdige algoritmer finnes, samt at det gir mening å forandre ett trinn om gangen. Mutasjoner utgjør fremmedelement i algoritmen, og vil forstyrre eller bryte opp en gitt algoritme.
vii) Mutasjoner forutsetter at ferdige algoritmer finnes, samt at det gir mening å forandre ett trinn om gangen. Mutasjoner utgjør fremmedelement i algoritmen, og vil forstyrre eller bryte opp en gitt algoritme.
viii) Naturlige utvalg forutsetter at virksomme algoritmer finnes, og kan ikke gjøre noe for å forandre dem. Naturlige utvalg kan ikke bygge algoritmer. Det kan bare komme inn og velge mellom ulike algoritmer, og velge noen bort ved å favorisere andre.
ix) Det finnes ingen entydige algoritmer i de grunnleggende fysiske naturlovene. Alle kjente lover for den døde natur ser ut til å være ikke-algoritmiske.
x) a) algoritmer kan være inkonsistente og ikke gjøre det de er ment å gjøre. b) algoritmer kan gå i stå eller stanse opp c) algoritmer kan gå i stykker
Darwinistenes såkalte mekanismer, er i utakt med algoritme-begrepet, f.eks. fordi mutasjoner som tilfeldig støy ikke er verken algoritmisk eller hierarkisk.
Homologi

Homologi er et samsvar mellom karakterer (egenskaper, organer osv.) hos forskjellige arter. Faktum er at hvis homologi defineres som likheter der skyldes felles avstamning, kan det ikke samtidig brukes som argument for samme felles avstamning. (En tar som utgangspunkt det en skal begrunne.) Hva enn årsaken kan være til at homologi opptrer mellom ulike dyreklasser, så har det i hvert fall ikke å gjøre med ensartede gener:
Tiltagende problem med 'livstreet'
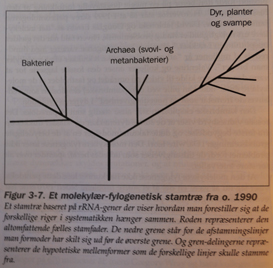 Moderne versjoner av det darwinistiske Livets stamtre, kalles "Fylogenetisk stamtre". På et typisk slikt tre, representerer "roten" stamformen til alle andre organismer i treet. De nederste greinene tegner de avstamningslinjer som forholdsvis tidlig har skilt seg ut, mens de høyere står for senere oppdelinger. Endene av grenene viser de egentlige arter. Oppdelingspunktet mellom to grener angir den hypotetiske stamformen for de to grenene. Mange slike trær tegnes slik at grenenes lengde svarer til forskjeller i sekvenser, som i sin tur skal indikere hvor lang tid det er gått siden de gikk hver til sitt (Fig3-7)
Moderne versjoner av det darwinistiske Livets stamtre, kalles "Fylogenetisk stamtre". På et typisk slikt tre, representerer "roten" stamformen til alle andre organismer i treet. De nederste greinene tegner de avstamningslinjer som forholdsvis tidlig har skilt seg ut, mens de høyere står for senere oppdelinger. Endene av grenene viser de egentlige arter. Oppdelingspunktet mellom to grener angir den hypotetiske stamformen for de to grenene. Mange slike trær tegnes slik at grenenes lengde svarer til forskjeller i sekvenser, som i sin tur skal indikere hvor lang tid det er gått siden de gikk hver til sitt (Fig3-7)
Med få unntak er de eneste 'riktige' data de som svarer til gren-endene på figuren. Sammensetningen av resten av treet er satt sammen ut fra metodologiske formodninger og sammenliknende sekvensanalyser. Det har vært en utbredt oppfatning om at jo flere molekyler som brukes i analysen, jo mer pålitelig må resultatet bli. Men denne forventningen begynte å smuldre opp for flere tiår siden. Molekylærbiolog James Lake m.fl. skrev i 1999 at "da forskere begynte analysere en bred vifte av gener, fra ulike organismer, fant de at organismenes innbyrdes forhold var i motstrid med rRNA-stamtreet". I følge de franske biologer Hervé Philippe og Patrick Forterre "viste det seg med flere og flere sekvensanalyser for hånden -at de fleste protein-fylogeneser er i motstrid både med hverandre og med mRNA stamtreet.
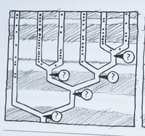 Med andre ord: ulike molekyler fører til høyst ulike fylogenetiske stamtrær. Biologen Carl Woese en av pionerene bak konstruksjonen av rRNA-baserte stamtrær, kommenterte: "Inntil videre har ingen troverdige organisme-fylogeneser sett dagens lys, ut fra de mange individuelle protein-fylogeneser en har laget. Brudd på den fylogenetiske kontinuitet forekommer overalt i stamtreet. Det mangler sammenheng mellom roten og oppover. De større forgreininger henger ikke sammen. Gruppene innbyrdes gjør det heller ikke. Og dannelsen av de grunnleggende grupperinger henger heller ikke sammen.
Med andre ord: ulike molekyler fører til høyst ulike fylogenetiske stamtrær. Biologen Carl Woese en av pionerene bak konstruksjonen av rRNA-baserte stamtrær, kommenterte: "Inntil videre har ingen troverdige organisme-fylogeneser sett dagens lys, ut fra de mange individuelle protein-fylogeneser en har laget. Brudd på den fylogenetiske kontinuitet forekommer overalt i stamtreet. Det mangler sammenheng mellom roten og oppover. De større forgreininger henger ikke sammen. Gruppene innbyrdes gjør det heller ikke. Og dannelsen av de grunnleggende grupperinger henger heller ikke sammen.
Woese tok hovedsakelig fatt på de uoverensstemmelser som befinner seg på systematikkens høyere nivåer, 'rikene'. Men problemet berører også mindre grener, herunder fylogenese for dyr. Biologen Michael Lynch skrev i 1999: "Å få klarhet i de større dyrerekkenes innbyrdes forbindelser har vist seg være et problem det er vanskelig å definere nærmere". Analyser av forskjellige gener, ja selv analyser av de samme gener har bragt en mengde ulike fylogenetiske stamtrær for dagen. Selv i de tilfeller der ulike molekylanalyser kan settes sammen, er resultatet ofte bisart.
F.eks. plasserer en studie fra 1996 som har anvendt 88 proteinsekvenser, kaniner i samme gruppe som primatene i stedet for hos gnagerne. En analyser fra 1998 plasserer kua i nærmere slektskap med hvalen enn med hesten. Den molekylære fylogenesen synes være blitt plassert i en sump med mudder til halsen.
Til tross for de nyere oppdagelsene innen molekylær fylogenese, holder National Academy of Scienses bastant fast ved at "molekylærbiologiens vitnesbyrd om evolusjonen som overveldende, og det vokser til stadighet" Det virker som de ikke holder følge med den forskning de selv skulle lede an i. Det blir da ikke så rart at lærebøker i biologi ikke følger opp, selv om 100 års-søvn er alt for lenge (se Haeckels fostre i min blogg). De tviler ikke et sekund på  "det overveldende vitnesbyrd i Livets Stamtre er en realitet, og at ingen forsker så mye som ville drømme om å tvile på en altomfattende felles avstamning."
"det overveldende vitnesbyrd i Livets Stamtre er en realitet, og at ingen forsker så mye som ville drømme om å tvile på en altomfattende felles avstamning."
Overganger mellom arter
Tidligere fantes en overtro på 'spontan generasjon'. Denne er ikke oppgitt, selvom den er motbevist av Pasteur, bare flyttet bakover i tid. Den samme unnvikelsesmanøveren brukes for alle betente overganger mellom artene på vei fra amøbe til menneske. Det er visstnok ingen eksisterende dyr som stammer fra andre typer konkrete dyr lenger. Mennesket stammer ikke lenger fra apene, men 'har felles apelignende forfedre med apene.' En flytter hele spekulasjonen om en felles avstamning akkurat passe langt tilbake i fortidas tåkeheim, til at det skal være umulig å motsi spekulasjonen.
Kan homologi-sirkelen brytes?
Tilsynelatende er det tre måter å unngå sirkeslutningen en kommer inn i om en både definerer og forklarer homologi ut fra felles avstamning. Den ene er å bruke homologi som definisjon på felles opphav, men unngå å utlede felles avstamning ut fra den. M.a.o. anerkjenne at homologi ikke lenger underbygger utviklingslæren. Evolusjonsbiolog David Wake skrev i 1999: Derfor "er homologi den forutsatte og forventede konsekvens av evolusjon. Homologi er ikke et vitnesbyrd om evolusjon."
Den andre måten er å gjeninnføre den pre-darwinistiske definisjon på homologi, som en strukturlikhet og samtidig anerkjenne at dette gjenåpner spørsmålet hvorvidt 'nedarving med tilpasning' er den beste  forklaring på fenomenet. I USA kan det være vanskelig å finne fortalere for denne retningen, fordi det blant biologer er meget upopulært og risikabelt profesjonelt å sette spørsmålstegn ved Darwinismen.
forklaring på fenomenet. I USA kan det være vanskelig å finne fortalere for denne retningen, fordi det blant biologer er meget upopulært og risikabelt profesjonelt å sette spørsmålstegn ved Darwinismen.
Den tredje og for tiden mest populære måte å løse problemet på er å definere homologi som uttrykk for felles avstamning, og derpå finne forskningsresultater som peker i retning av nedarving med tilpasning, uavhengig av homologien. Slike forskningsresultat kan være strukturorientert, f.eks. i form av DNA-sekvenssamlinger eller fossilmaterialet. Eller det kan være prosessorientert: utviklingsretninger eller utviklingsgenetikk. De to første legger ut med en antagelse om felles avstamning og førsøker dernest å utlede det mest sannsynlige mønster for de ulike slektskapsforhold. De to sistnevnte forsøker finne de prosesser som kan være ansvarlige for likheter som skyldes felles avstamning.
Hva med fossiler?
 Noen biologer har hevdet at en måte å fastsette evolusjonære avstamningsforhold på, er å spore likhetene i to eller flere organismer via en ubrutt kjede av fossile organismer, tilbake til sitt fossile opphav. Uheldigvis er det å sammenligne fossiler ikke mindre vanskelig enn å sammenligne levende prøveeksemplarer. Sokal og Sneath skrev i 1963: "Selv om det er fossilmateriale tilgjengelig, må dette materiale først fortolkes" ved sammenlikning av ensartede trekk. Ethvert forsøk på å dra slutninger om
Noen biologer har hevdet at en måte å fastsette evolusjonære avstamningsforhold på, er å spore likhetene i to eller flere organismer via en ubrutt kjede av fossile organismer, tilbake til sitt fossile opphav. Uheldigvis er det å sammenligne fossiler ikke mindre vanskelig enn å sammenligne levende prøveeksemplarer. Sokal og Sneath skrev i 1963: "Selv om det er fossilmateriale tilgjengelig, må dette materiale først fortolkes" ved sammenlikning av ensartede trekk. Ethvert forsøk på å dra slutninger om  fossilenes evolusjonære slektskap som bygger på homologi som begrunnelse for felles avstamning " fanger oss hurtig inn i et nett av sirkelslutninger som det er umulig å unnslippe."
fossilenes evolusjonære slektskap som bygger på homologi som begrunnelse for felles avstamning " fanger oss hurtig inn i et nett av sirkelslutninger som det er umulig å unnslippe."
Faktisk er det vanskeligere å utlede evolusjonært slektskap ut fra den fossile lagserie enn fra levende organismer, fordi fossilhistorien er fragmentarisk, og fossilene ikke har bevart alle relevante trekk. Biologen Bruce Yong skrev i 1993: "Hvis fossiler overhodet er noe, så er de i hvert fall av mindre verdi når man skal fastslå homologe forhold, ettersom de normalt omfatter langt færre egenskaper enn levende organismer."
Men selv om den fossile lagserie hadde vært fullstendig, og den hadde bevart alle ønskede egenskaper, ville den likevel ikke godtgjøre at homologi skyldes et felles opphav. Det finnes andre forklaringer enn nedarving ved tilpasning på morfologiske likheter. De fleste biologer før Darwin ville ha forklart slike likheter med felles design eller skapelse. Fossilmaterialet er forenlig med begge forklaringer. En enkel oppstilling av former som ligner hverandre, gir ikke en forklaring på opphavet i seg selv. Det kreves en mekanisme som kan forklare slike likheter. Forklaringer som "nedstamning og tilpasning" er bare ord, om de ikke kan kobles mot faktiske biologiske prosesser.
Darwinistisk mytologi følger et ullenhetsprinsipp i sin skjødesløse omgang med fortida. Den benekter at det fantes skarpe grenser mellom livsformer. En hevder på en utydelig måte ('den røde dronning') at alle livsformer langt nok tilbake i tid 'fløt over i hverandre'. Vi kan øyne tre darwinistiske prinsipper midt oppi all utydeligheten:
i) 'Livsformene var tidligere mer utydelige og primitive og mindre differensierte enn de er i dag.' ii) 'Det eksisterer ingen grense mellom algoritmiske og ikke-algoritmisk liv.' iii) 'Overlevelse var aldri noen problemstilling'. Før i tiden var 'alt så mye bedre': en hadde liksom livsformer som både var encellet og flercellet samtidig; var litt bløtdyr og litt virveldyr samtidig, og litt krypdyr og litt pattedyr samtidig. Det er til tross for de enorme forskjeller mellom krypdyr og pattedyr. Her er noen av de vesentligste anatomiske forskjellene:
 a) Krypdyr legger egg, pattedyr har livmor og føder levende unger. b) Krypdyr har eggeplomme, pattedyr har melkekjertler. c) Krypdyr har kloakkåpning, Pattedyr har separate kroppsåpninger for forplantning og avføring. d) Krypdyr har skjell, pattedyr har hud med hår. e) Krypdyr har fem kjeveledd på hver side, pattedyr har bare ett kjevebein på hver side. f) Krypdyr har primitiv hørsel, pattedyr har øreknokler som gir bedre hørselsbilde. g) Krypdyr er vekselvarme, pattedyr er varmblodige.
a) Krypdyr legger egg, pattedyr har livmor og føder levende unger. b) Krypdyr har eggeplomme, pattedyr har melkekjertler. c) Krypdyr har kloakkåpning, Pattedyr har separate kroppsåpninger for forplantning og avføring. d) Krypdyr har skjell, pattedyr har hud med hår. e) Krypdyr har fem kjeveledd på hver side, pattedyr har bare ett kjevebein på hver side. f) Krypdyr har primitiv hørsel, pattedyr har øreknokler som gir bedre hørselsbilde. g) Krypdyr er vekselvarme, pattedyr er varmblodige.
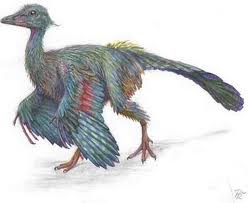
Til sammenligning kan vi tenke på urfuglen (Archaeoteryx), og dens status som mellomform mellom krypdyr og fugler. Den var uten tvil en fugl: den hadde fjær og ikke skjell. Den hadde nebb og ingen kjeve. Den var ganske sikkert varmblodig, og ikke vekselvarm. Den hadde vinger med fjær, som den flakset med når den fløy. Intet krypdyr har hatt fjær eller fuglevinger. Flyveøgler hadde svevehud, i likhet med flaggermus. Det var to ting som fikk 'urfuglen' til å ligne på et krypdyr: Det ene var at den hadde masse små tenner. Det andre at den hadde klør på vingene. Til tross for sine små likheter med krypdyr, har Archaeoteryx en ikonisk status i darwinismen. Den betraktes som et 'endegyldig bevis' på overgangen mellom krypdyr og fugler. Den visker liksom ut alle grenser og gjør overgangen mellom krypdyr og fugler til en 'naturlig og hverdagslig' sak.
Lunger hos krypdyr og fugler
I følge evolusjonsteorien skal fugler komme fra krypdyr. Ett av de mange problemene de da støter på er lungesystemets overgang, fra et motstrømssystem hos krypdyr -likt med pattedyr, til ett sirkulært system som hos fugler.
Bilde. 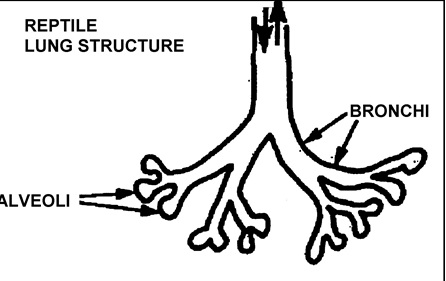 Lungestruktur hos krypdyr
Lungestruktur hos krypdyr
 Krypdyr har i prinsippet samme type lunger som pattedyr (utskifting via alveoler). Da returnerer luft samme vei som den kom inn i lungene, via bronkiene, for deretter å bli utskilt til blodårene via alveolene.
Krypdyr har i prinsippet samme type lunger som pattedyr (utskifting via alveoler). Da returnerer luft samme vei som den kom inn i lungene, via bronkiene, for deretter å bli utskilt til blodårene via alveolene.
Bilde 2. Menneskers lungesystem
Hos fugler er lungesystemet totalt annerledes tilvirket: hentet fra Ask Nature. Når frisk luft først blir inhalert gjennom fuglesvelget (neseborene), reiser den gjennom luftrøret (et stort rør som strekker seg fra halsen), som deler seg i venstre og høyre primære bronkier (aka mesobronchi, med hver bronkie fører til en lunge). Inhalert luft reiser ned hver primær bronkie og deretter deler den seg: litt luft inn i lungene der gassutvekslingen skjer, mens den resterende luften fyller posterior (bakre) luftblærer. Deretter, i løpet av første utpust, går den friske luften fra de bakre sekker inn i lungene, og gjennomgår så gassutveksling. Den brukte luften i lungene fortrenges av denne innkommende luft, og strømmer ut av kroppen gjennom luftrøret.
bronkie fører til en lunge). Inhalert luft reiser ned hver primær bronkie og deretter deler den seg: litt luft inn i lungene der gassutvekslingen skjer, mens den resterende luften fyller posterior (bakre) luftblærer. Deretter, i løpet av første utpust, går den friske luften fra de bakre sekker inn i lungene, og gjennomgår så gassutveksling. Den brukte luften i lungene fortrenges av denne innkommende luft, og strømmer ut av kroppen gjennom luftrøret.
Bilde. Prinsippskisse av fuglers respirasjon
Under den andre innånding, kommer igjen frisk luft inn gjennom både de bakre sekker og lungene. Forbrukt luft i lungene blir igjen fortrenges av innkommende luft, men det kan ikke gå ut gjennom luftrøret fordi frisk luft strømmer innover. I stedet går brukt luft fra lungene, til fremre luftblærer. Deretter, i løpet av den andre utånding, strømmer den brukte luften i fremre blærer og i lungene, ut gjennom luftrøret, og frisk luft i de bakre sekkene inn i lungene for gassutveksling.
 Dette mønsteret for luftstrøm gjennom luftveiene, skaper enveis strøm av frisk luft i løpet av gassutvekslings-flatene i lungene. Videre passerer frisk luft gjennom gassutvekslingsflatene under både inhalering og utånding, noe som resulterer i en konstant tilførsel av frisk luft slik at fuglen settes i stand til å oppleve en nesten kontinuerlig tilstand av gassutveksling i lungene.
Dette mønsteret for luftstrøm gjennom luftveiene, skaper enveis strøm av frisk luft i løpet av gassutvekslings-flatene i lungene. Videre passerer frisk luft gjennom gassutvekslingsflatene under både inhalering og utånding, noe som resulterer i en konstant tilførsel av frisk luft slik at fuglen settes i stand til å oppleve en nesten kontinuerlig tilstand av gassutveksling i lungene.
Bilde. Fugler har behov for konstant luftgjennomstrømning
Dette i motsetning til krypdyrs lunger, som altså har toveis luftstrømmer over gassvekslingsoverflatene. Bilde. Lungenes lveoler -utveksling av oksygen med blodet:
 Darwinister prøver få seg selv og andre til å akseptere at alle skarpe grenser, er noe naturen har funnet på senere! Vesener som er litt det ene og litt det andre, er 'ulne': En ullen livsform uten presise spesifikasjoner, har ikke mer enn en 'liksom-overlevelse'. Om dette gjelder de fleste livsformer i omgivelsene, overser man en helt avgjørende sak: 'Å være levedyktig er viktigere enn å hevde seg i konkurransen.' Fordi konkurransen visstnok var så svak 'en gang i fortiden', ser en helt bort fra alle fint avstemte algoritmiske mekanismer som må samvirke, om et individ i det hele tatt skal klare seg. Overlevelse synes være et lite problem for dem, i forhold til det å konkurrere med de andre livsformene. Men naturlig utvalg velger, som før nevnt, bare bort ting. Når en velger noe, velger en samtidig vekk noe annet. En som skal gifte seg, velger éi dame og samtidig bort de andre..
Darwinister prøver få seg selv og andre til å akseptere at alle skarpe grenser, er noe naturen har funnet på senere! Vesener som er litt det ene og litt det andre, er 'ulne': En ullen livsform uten presise spesifikasjoner, har ikke mer enn en 'liksom-overlevelse'. Om dette gjelder de fleste livsformer i omgivelsene, overser man en helt avgjørende sak: 'Å være levedyktig er viktigere enn å hevde seg i konkurransen.' Fordi konkurransen visstnok var så svak 'en gang i fortiden', ser en helt bort fra alle fint avstemte algoritmiske mekanismer som må samvirke, om et individ i det hele tatt skal klare seg. Overlevelse synes være et lite problem for dem, i forhold til det å konkurrere med de andre livsformene. Men naturlig utvalg velger, som før nevnt, bare bort ting. Når en velger noe, velger en samtidig vekk noe annet. En som skal gifte seg, velger éi dame og samtidig bort de andre..
Felles avstamning -for hvem?
Harry Whittington, som var paleontologen som først åpenbarte omfanget av den kambriske eksplosjonen, skrev i 1985: "Jeg ser med stor skepsis på de diagrammer som viser at dyrelivets diversitet skulle være resultat av en forgrening opp gjennom tiden, og som koker det ned til avstamning fra en eller annen form for dyr. . Dyrene kan meget vel være  oppstått mer enn én gang og på ulike steder til ulike tider." Etter å ha fått tilgang til nyeste informasjon om molekylær fylogenese, skriver biologen Malcolm Gordon i 1998: "livet ser ut til å ha hatt atskillige opprinnelseskilder. Det universelle livets stamtre synes å ha mer enn bare en rot.. Den tradisjonelle versjon av teorien om felles avstamning, gjelder tilsynelatende ikke for riker ..[eller] rekker, og muligens heller ikke for mange av klassene innenfor rekkene.
oppstått mer enn én gang og på ulike steder til ulike tider." Etter å ha fått tilgang til nyeste informasjon om molekylær fylogenese, skriver biologen Malcolm Gordon i 1998: "livet ser ut til å ha hatt atskillige opprinnelseskilder. Det universelle livets stamtre synes å ha mer enn bare en rot.. Den tradisjonelle versjon av teorien om felles avstamning, gjelder tilsynelatende ikke for riker ..[eller] rekker, og muligens heller ikke for mange av klassene innenfor rekkene.
Kompetente biologer setter spørsmålstegn ved det darwinistiske Livets Stamtre. Bedømt ut fra det fossile og molekylære vitnesbyrd, har vi her ikke å gjøre med en velbegrunnet teori. Men om en reiser spørsmål ved det, blir det vanligvis ikke sett på med milde øyne. Særlig gjelder det kanskje USA, men også her i Norge kan en merke motstanden, -også på kirkelig hold. Det er fullt tillatt å kritisere styrende i stat og kirke, men å kritisere Darwin blir ikke vel mottatt..
Juks med rekapitulasjonsteorien
Haeckels biogenetiske lov hadde stor oppslutning før og etter år 1900, men så mistet den oppslutning i 1920 årene. Stephen Jay Gould mener at 'den biogenetiske lov først falt i unåde da den ble umoderne'. Vitenskapshistorikeren Nikolas Rasmussen er enig: 'Alt det forskningsmateriale som skal til for å forkaste den biogenetiske lov har vært til stede fra den første dag loven ble akseptert'. Det var helt åpenbart at en ikke gikk vekk fra den fordi nye oppdagelser motsatt den, men at den med ett ble 'umoderne'.
Til tross for at rekapitulasjonsteorien 'gikk av moten', forsøkte enkelte forskere (bl. a. Lillie) holde deler av den i live av 'tilpasnings grunner' i forhold til evolusjonsteorien. Men i 1922 ble den holdt for å være 'demonstrativt utroverdig' av den britiske embryolog Walter Garstang. Det var fordi 'de ontogenetiske trekk ikke bar på et eneste vitnesbyrd om de fullvoksne trekk som hører avstamningen til'. Garstang hadde ingen tillit til Haeckels hypotese om at nyutviklede trekk hekter seg som nye ledd på kjeden i fosterutviklins-prosessen: 'En hytte blir ikke et hus bare fordi en setter på en ekstra etasje ovenpå. Et hus representerer en høyere posisjon innen for boligutvikling, men det krever samtidig at hele bygningen bygges om.' (dette må tolkes ut fra 1922 da hytter framdeles var hytter.)
Garstang visste altså at 'den biogenetiske lov' ikke passer med forskningsmaterialet. Men på grunn av deres tro på darwinistisk evolusjon, var de overbevist om at en eller annen form for rekapitulasjon måtte være sann. Embryologen Gavin de Beer ga i årene 1940 til 1958 ut tre utgaver av en bok om embryologi og evolusjon, der han kritiserer Haeckels 'biogenetiske lov'. Her sier han bl.a.: 'Med andre ord utviser de tidlige utviklingsstadier avgjørende forskjeller (hos ulike arter-min tilføyelse)..og det i motsetning til Darwins tro på at de er meget like. De Beer konkluderte med at rekapitulasjon er en mental tvangstrøye, som har motarbeidet og forsinket utviklingen i embryologi'.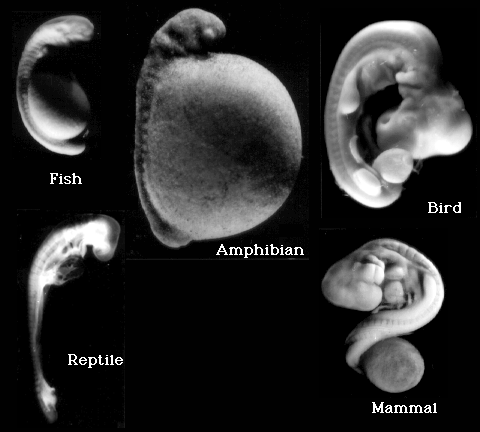
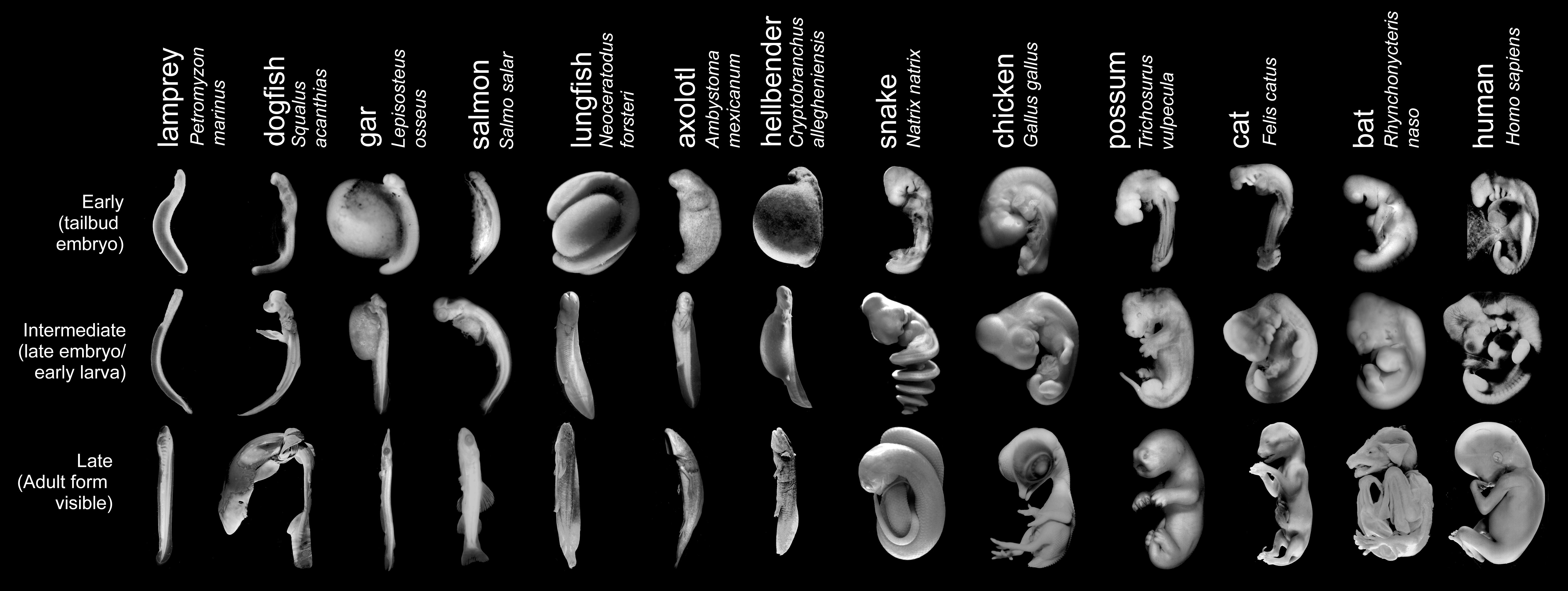
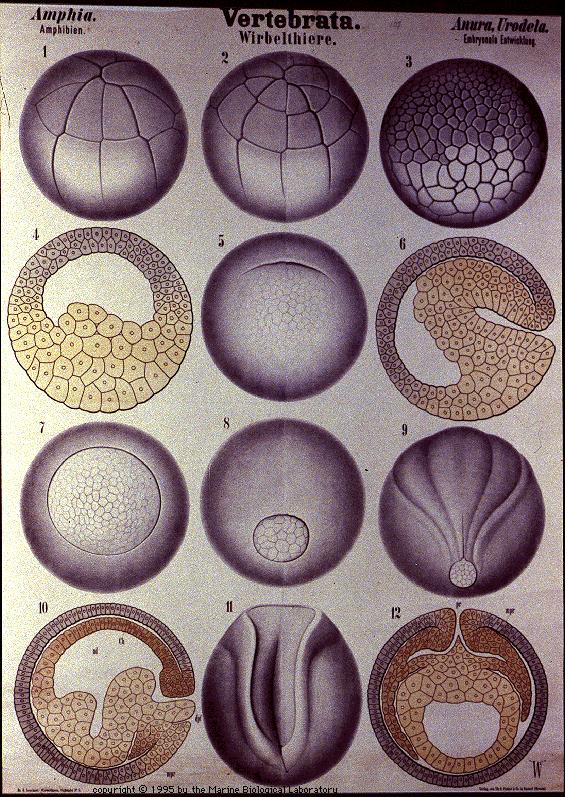
Tidlige fostres ulikheter er velkjente
Biologer har i over 100 år kjent til tidlige virveldyr-fostres ulikheter. Embryologen Adam Sedgwick understreket i 1894 at 'von Baers lov' om tidligere likheter ..ikke stemmer overens med det en kan slå fast om fosterutviklingen. Sedgwick sammenlignet fosteret av en liten hai med det til en høne og slo fast: 'Det finnes intet utviklingsstadium, hvorpå en så lett som ingenting kan se forskjell på dem med det blotte øye'.. 'Hvis von Baers lov skal gi mening, må den forutsette at dyr så nærbeslektet som høns og ender, ikke er til å skjelne fra hverandre på de tidligste stadier.. Ikke desto mindre kan jeg se forskjell på et høne og ande-embryo fra dag to" I følge Sedgwick kan "en art skjelnes fra alle de andre fra fosterutviklingens første stadier og hele veien opp." I det 19.århundre kritiserte von Baer darwinistene for overhodet ikke ha undersøkt fostrene.
Embryologer fra nåtiden bekrefter dette. I 1997 bemerket Erich Blechschmidt: "De tidligste stadier i menneskets fosterutvikling atskiller seg fra alle andre arters." Og i 1987 rapporterte Richard Elingson at frø, høns og mus er "vesensforskjellige når det gjelder fundamentale egenskaper som egg-størrelse, befruktningsmekanismer, kløvningsmønstre og [gastrukulære] bevegelser".
Etter å ha utviklet seg i ulike retninger på tidligere stadier, blir virveldyrfostre overraskende nok litt mer like hverandre ca. halvveis i deres utvikling. Det er dette midtveis-stadium Haeckel anvendte som sitt 'første' stadium i sine tegninger. Skjønt han overdrev disse likhetene vidt og bredt, er det likevel -i hvert for amatører -visse likheter til stede.
I 1976 kalte William Ballard det 'faryngula' på grunn av de 'kammer/poser' som opptrer parvis på begge sider av spiserøret. I følge Richardson er det viktig at hos virveldyr utvikler de (i det skjulte) kroppsstrukturens legemstrekk over en lang rekke stadier, ikke bare på ett som noen hevder. For amatører synes det likevel tilsynelatende som utviklingsforløpet av fostre har form av et 'utviklingstimeglass' Han bemerker at von Baers lov gir en ufullstendig beskrivelse. Han beskjeftiger seg faktisk kun med den siste halvdel av ontogenesen(det enkelte individs embryoiske utvikling).
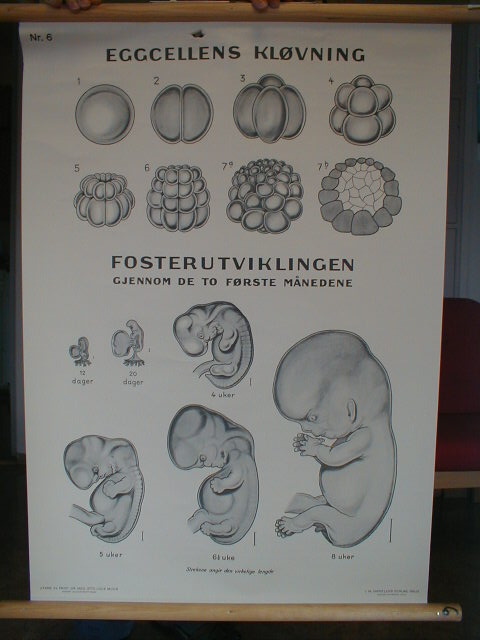
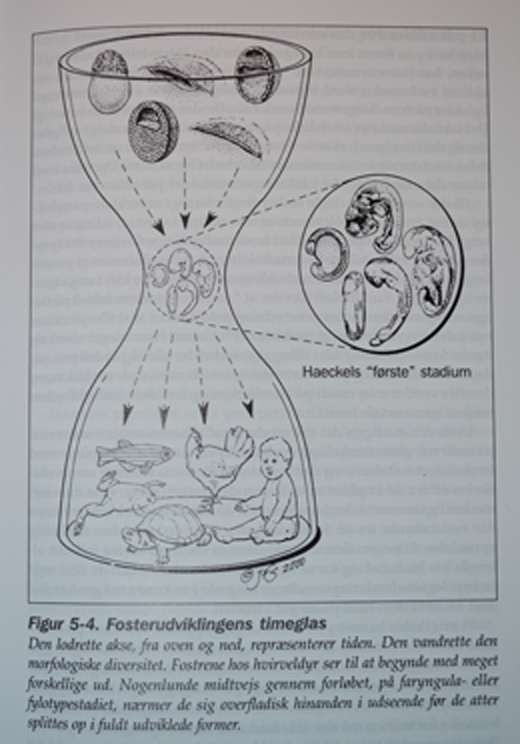 Et paradoks for darwinistisk utviklingsteori
Et paradoks for darwinistisk utviklingsteori
Hvis det er slik at von Baers lov kun gjelder for siste halvdel av ontogenesen, har Darwin mistet sitt aller viktigste enkeltargument for sin idé om nedstamning med tilpasning. I følge Darwin var det fostrenes likheter på deres tidligste stadier som skulle vise den felles avstamning. Det faktiske mønster: 'først ulikhet -så mere likheter- så mer ulikheter igjen', forekommer temmelig uventet i forbindelse med darwinistisk evolusjon. Så i stedet for å støtte Darwins teori, utgjør det embryologiske forskningsmateriale et paradoks for den.
I det senere har embryologer forsøkt tilpasse dette til darwinistisk teori, ved å snakke om 'stor frihet i variasjonen i starten når det skal dannes nye utviklingsmønstre og livshistorier..' Uansett hvor mye vekt slike 'fortellinger' skal tillegges, står det fast at de baserer seg på en formodning om darwinistisk evolusjon. Så 'vender man rundt på tallerkenen' og fortolker embryologiens forskningsmateriale ut fra dette. Det blir jo det motsatte av å basere evolusjonsteorien på forskningsresultater. Hvis man gjør det, vil man formodentlig komme til at de ulike virveldyrklasser ikke har en felles stamfar. For naturalistisk darwinisme, blir dette uspiselig. En blir nødt til å gjenfortolke resultatene for å få dem til å passe inn i teorien.
I det 19.århundre kritiserte von Baer darwinistene for overhodet ikke ha undersøkt fostrene. Det er ikke så mye annerledes i dag. Det virker likegyldig for mange hvor mye embryologien er i uoverensstemmelse med evolusjonsteorien. For teorien må en åpenbart ikke sette spørsmålstegn ved. Til tross for bevislig falske tegninger og at Haeckels biogenetiske lov gjentatte ganger er tilbakevist, fortsetter de å dukke opp igjen og igjen.
Lærebøker i biologi fortsetter å bringe Haeckels falsknerier til torvs. Og Haeckels biogenetiske lov: 'Ontogenesen rekapitulerer fylogenesen' (rekapitulasjonsteorien), framsettes i stadig nye versjoner. Vi tar ikke med alle de bøker det er snakk om her, men som følge av denne hyppige feilen gjentar vi de alvorligste problemene ved den:
1: Haeckels tegninger tar utelukkende med de klasser og ordener som passer best med hans teori.
2. Tegningene forvansker/tillemper de fostre de påstår å vise -og mest alvorlig:
3. De utelater fullstendig de stadier der virveldyrfostre er ytterst forskjellige!
 Er et menneskefoster likt fisk?
Er et menneskefoster likt fisk?
Det er ikke bare i lærebøkene at foto av fostre forleder leseren til å tro på rekapitulasjon. Et eks. er i november utgaven av Life (1996). Den inneholder foto av et menneske, en ape, en lemur, en gris og en kylling -alle på fosterstadiet. Biolog Kenneth Miller (Brown University) avviser rekapitulasjonsteorien, som et eks. på at 'skinnet kan bedra selv en dyktig forsker'. Likevel beskriver Miller hvordan mennesket kan utvikle ..noe som i høy grad ligner gjeller. Men det kommer an på øynene som ser. Om en tolker bildene inn i Haeckels biogenetiske lov, så kan en komme til slikt resultat. Om en ikke gjør det, kan en komme til et helt annet.
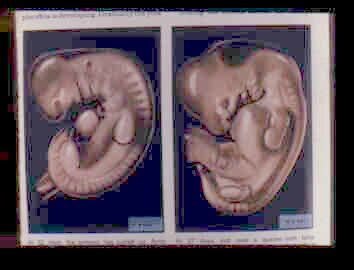 'Gjeller' er ikke gjeller.
'Gjeller' er ikke gjeller.
Halvveis i fosterutviklingen har alle virveldyrfostre en rekke folder i området ved halsen (pharynx). Foldenes utadvendte deler har et 'fure'lignende utseende, og er ikke gjeller. De er ikke engang gjeller i starten hos fisk inni egget. De utvikles først senere. Hos krypdyr, fugler og pattedyr utvikler de seg til å bli helt andre strukturer, slik som det indre øret og skjoldbruskkjertler. Den tilsynelatende likheten skyldesnoen overfladiske furer i halsregionen. I følge den britiske embryologen Lewis Wolpert er 'denne likheten illusorisk. Den ligner alene de strukturer en fisk har på fosterstadiet, som gir opphav til gjeller'.
Eneste måte å kunne tale om gjellelignende strukturer hos menneskefostre, er ved å lese evolusjon inn i fosterutviklingen. Men har en gjort det, kan en ikke så bruke fosterutvikling til å bekrefte evolusjonsteorien. Det er et typisk sirkelargument, som også benyttes angående homologi til felles opphav. Tross forsikringer om at ingen lenger tror på rekapitulasjons-teorien, gjør folk det likevel. At gjeller ikke hører med til fosterstrukturen, ikke engang hos fisk, er ukjent for mange. I stedet får en ofte høre: 'se hvor lik de er på fosterstadiet'. At en 'ser' gjeller på fosterstadiet, er imidlertid å lese en struktur inn i fosteret.
Utvalg og bilder ved Asbjørn E. Lund