 Parafrase til innledning av boka Zombi-science
Parafrase til innledning av boka Zombi-science
(Kursivering og understreking ved oversetter.)
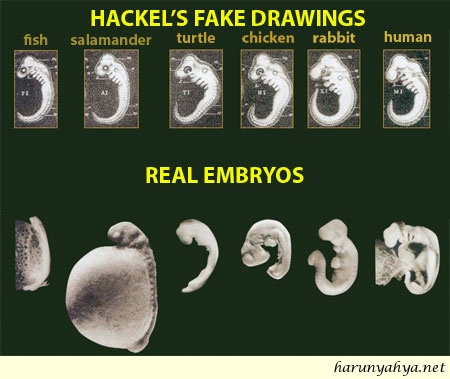
I 2000 kom J. Wells med en bok 'Icons of Evolution'-her; Den inneholdt viktige temaer som evolusjon tidligere var markedsført under. Wells viser der at disse punktene enten er feilaktige (eks. Haeckels fostre) og/eller ikke indikerer det de er ment å gjøre (eks: nattsvermere). Til tross for en grundig tilbakevisning av påstandenen i disse, er lite skjedd. Riktignok har visstnok én lærebokforfatter endret på en bevisst fordreining fra tidligere, men i det store og hele har det vært 'same procedure as usual'. Dette gjelder også i forhold til hans 2. bok i 2011 om 'Junk DNA'. Her har undertegnede selv opplevd i avis-diskusjon at det har vært hevdet fra evolusjonistisk hold, at det ikke er noen motsetning mellom det som nå er funnet at mer enn 80% av DNA har en funksjon i forhold til tidligere påstander fra evolusjonistisk hold om at mesteparten av menneskelig DNA er overlevninger (rest-DNA) fra tidligere.
Bilde 1. Omslagsbilde i Wells' 1. bok
Grunnen til at Wells benytter uttrykket 'zombi-science' er at det selv etter å ha blitt påvist klare feil, fortsatt, med uforminsket energi, benyttes samme argumenter fra darwinister. Dette har historiske røtter: På slutten av 1800-tallet hang de fast i LaMarcks arvelære. Det var først etter flere tiår at de på 1900-tallet gikk de over til Mendels posisjon m.h.t. arvelære. Metodologisk naturalisme var rådende, og en var på jakt etter molekyler som utgjorde arvemterialet. I løpet av 1.halvdel av 1900-tallet dreide forskningen fra proteiner til DNA. Det ble kalt 'the transforming principle' ('det forvandlende prinsipp'), og jakten på DNA's oppbygning førte fram til Watson og Cricks modell for DNA som en dobbel vindeltrapp, bestående av et par av maler (templates), som er komplementære til hverandre.
I 1958 foreslo Fr. Crick at det spesifikke ved et DNA-segment ligger i sekvensen av nukleotider, som den er bygd opp av. Det sentrale dogmet i biologien er knyttet til denne sekvensen av nukleotider, samt at DNA-sekvenser kan overføres til proteiner. I 1970 skrev Jacob at organismen er realiseringen av 'et genetisk program' skrevet i DNA-sekvenser. (20) Dermed syntes informasjon å kunne reduseres til materie, og sentral-dogmet ble materiell basis for arv, utvikling og evolusjon. En lærebok gikk så langt at den hevdet at 'evolusjon kan presist defineres som enhver endring i frekvensen til alleller innen en gen-pool fra en generasjon til den neste. (23)
Bilde 2. Omslagsbilde i Wells' 3. bok
Ikke alle var enige i det. I 1963 kalte Ernst Mayr tilnærmingen ovenfor 'bønne-genetikk' (beanbag genetics), fordi den betraktet gener som usammenhengende i stedet for samvirkende. (24) Mayr skrev at 'beanbag genetics' på mange måter er villedende, fordi ett individ er ikke en mosaikk av karaktertrekk, som alle er produkt av ett gitt gen. Gener er heller enhetene i et genetisk program som styrer den kompliserte utviklingsprosessen." (25) Sean B. Carroll støttet Mayrs kritikk (2005): "utvikling av form er hoveddramaet i livets historie.. Så la oss lære bort den historien: I stedet for 'endring i gen-frekvensen, la oss forsøke evolusjon av form er endring i utviklingen." (26)
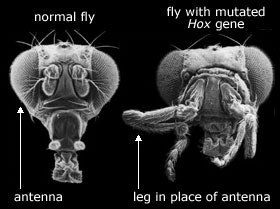
 I 1992 skrev biologen Brian K. Hall at til tross for "dominansen i evolusjon av populasjons-genetikk," er det "verken tilstrekkelig eller inklusivt." Spesielt ignorerer populasjons-genetikk embryo-utvikling. Hall foreslo en ny disiplin som han kalte evolusjonær utviklings biologi (evo-devo). Et resultat av denne var fokusering på Hox-gener, som påvirker utvikling fra hode til hale, og har likhetstrekk innen ulike arter. Spørsmålet ble da: 'Hvis dyr har lignende utviklings-gener, hvorfor utvikler de seg så ulikt? I 1990 benyttet utviklingsbiolog Eric Davidson termen 'Gen-regulerende Nettverk' (GRN) for å betegne ett sett av samvirkende DNA-sekvenser, RNA og proteiner som regulerte oversettelsen (transkripsjonen). Konklusjonen var at dyr med underliggende lignende gener utvikler seg ulikt, grunnet forskjeller i hvor og når disse genene skrives av. Men Brian Hall hadde i 1992 skrevet at "forståelse av kontroll og utvikling vil kreve å bestemme hvordan genetiske, epigenetiske og omgivelsesfaktorer er integrert til et hierarkisk sett av samvirkende kontroller. En forståelse av genomet alene, vil selv om den er viktig, ikke være forklaringen vi søker. (32)
I 1992 skrev biologen Brian K. Hall at til tross for "dominansen i evolusjon av populasjons-genetikk," er det "verken tilstrekkelig eller inklusivt." Spesielt ignorerer populasjons-genetikk embryo-utvikling. Hall foreslo en ny disiplin som han kalte evolusjonær utviklings biologi (evo-devo). Et resultat av denne var fokusering på Hox-gener, som påvirker utvikling fra hode til hale, og har likhetstrekk innen ulike arter. Spørsmålet ble da: 'Hvis dyr har lignende utviklings-gener, hvorfor utvikler de seg så ulikt? I 1990 benyttet utviklingsbiolog Eric Davidson termen 'Gen-regulerende Nettverk' (GRN) for å betegne ett sett av samvirkende DNA-sekvenser, RNA og proteiner som regulerte oversettelsen (transkripsjonen). Konklusjonen var at dyr med underliggende lignende gener utvikler seg ulikt, grunnet forskjeller i hvor og når disse genene skrives av. Men Brian Hall hadde i 1992 skrevet at "forståelse av kontroll og utvikling vil kreve å bestemme hvordan genetiske, epigenetiske og omgivelsesfaktorer er integrert til et hierarkisk sett av samvirkende kontroller. En forståelse av genomet alene, vil selv om den er viktig, ikke være forklaringen vi søker. (32)
Bilde 3. Wells avslørte falske ikoner i tidligere bok
Epigenetikk (fra kap. 4)
 Epi (gresk) betyr over/i tillegg (meta-betydning). Ordet hadde flere betydninger fra starten: en smal betydning der det refererte til mekanismene som gener produserte fenotype-effekter. En bredere betydning omfattet alle faktorene involvert i utvikling, hvorav genomet bare er én slik. De fleste biologer benytter epigenetikk i dens smale betydning, 'arvelige kromosom-endringer som ikke endrer underliggende DNA-struktur'. Slike epigenetiske endringer påvirker når og hvor spesifikke DNA-sekvenser skrives av, til RNA. Biologi-historiker Jan Sapp har dokumentert hvordan gen-sentrert tenkning har blitt dominerende i løpet av det 20. århundre, men bevis har nå akkumulert seg om at et slikt trangt syn bryter med realitetene. Ved å gjenkjenne at det er epigenetiske arvelighetssystem som overfører ikke-DNA variasjoner i celler og organisme-utvikling, utvider arvelighetsbegrepet og utfordrer den vidt aksepterte gen-sentrerte neo-darwinistiske versjon av Darwinismen. (37)
Epi (gresk) betyr over/i tillegg (meta-betydning). Ordet hadde flere betydninger fra starten: en smal betydning der det refererte til mekanismene som gener produserte fenotype-effekter. En bredere betydning omfattet alle faktorene involvert i utvikling, hvorav genomet bare er én slik. De fleste biologer benytter epigenetikk i dens smale betydning, 'arvelige kromosom-endringer som ikke endrer underliggende DNA-struktur'. Slike epigenetiske endringer påvirker når og hvor spesifikke DNA-sekvenser skrives av, til RNA. Biologi-historiker Jan Sapp har dokumentert hvordan gen-sentrert tenkning har blitt dominerende i løpet av det 20. århundre, men bevis har nå akkumulert seg om at et slikt trangt syn bryter med realitetene. Ved å gjenkjenne at det er epigenetiske arvelighetssystem som overfører ikke-DNA variasjoner i celler og organisme-utvikling, utvider arvelighetsbegrepet og utfordrer den vidt aksepterte gen-sentrerte neo-darwinistiske versjon av Darwinismen. (37)
Noen biologer forstår epigenetikk bredere. I 1993 skrev Susan Herring: "Grovt sagt refererer epigenetikk til hele settet av samhandling mellom celler og celle-produkter", som fører til embryo-utviklingen. (36) I 2002 skrev Eva Jablonka og Marion Lamb at epigenetikk er "hovedsakelig opptatt av mekanismene som styrer celler til en spesiell form eller funksjon.. Når en gjenkjenner at det er epigenetiske systemer for arv, hvor ikke-DNA variasjoner kan bli overført i celle og organisme-linjer, utvider arv-begrepet og utfordrer den vidt-aksepterte gen-sentrerte neo-darwinistiske versjon av darwinismen." I denne bredere betydning innebærer den at arv og utvikling angår mye mer enn DNA. En må skille mellom å 'lage' og 'fullt spesifiserere': DNA spesifiserer ikke fullt ut RNA, RNA spesifiserer ikke protein fullt ut og protein spesifiserer ikke oss fullt ut.
Bilde 4. Epigenetisk tagging
Det er flere grunner til at DNA ikke fullt ut bestemmer RNA, som igjen bestemmer protein som ikke fullt ut spesifiserer oss. Én slik grunn er at når DNA-sekvenser blir oversatt til RNA, så blir mange RNA endret, slik at de ikke passer den opprinnelige 'oppskriften'. En måte det skjer på er ved 'alternativ spleising'. De fleste DNA-sekvenser i plante-og dyreceller, blir avbrutt av såkalte 'introner'. Disse settes inn når DNA oversettes til mRNA, men så spleises de protein-kodende regioner av DNA sammen, uten at disse intronene tas med. Spleisingen kan utføres på ulike måter, slik at mange av de endelige mRNA har sekvenser som ikke lenger svarer til DNA-sekvenser. F.eks. kan RNA fra én DNA-sekvens i bananfluer generere mer enn 18.000 ulike proteiner gjennom alternativ spleising. (39) Det gjelder også for mennesker. (40) Så påstanden at "DNA fullt spesifiserer RNA, og RNA fullt spesifiserer proteiner" er feilaktig, men det er i å danne oss, at sentral-dogmet til neo-darwinister feiler mest dramatisk.
 I tillegg til alternativ spleising kan DNA-oversetting hos mange dyr gjennomgå RNA-redigering -her, som kan endre aminosyre-sekvensen, og dermed funksjon til resulterende protein. F.eks. er det i blekkspruter funnet omfattende RNA-redigering for å tilpasse seg omgivelser, f.eks. kulde -her (41). Nylige studier har også avdekket omfattende RNA-redigering i mennesker. (42) For proteiner gjelder at selv om deres funksjon avhenger av deres tre-dimensjonale form, så bestemmes ikke alltid formen av aminosyre-sekvensen. Noen proteiner tilpasser lignende foldede former, til tross for meget ulike aminosyre-sekvenser. (43) Andre antar ulike former, til tross for at de har samme eller meget like aminosyre-sekvenser. Det siste er kjent som 'forvandlings' (metamorphic) proteiner. (44)
I tillegg til alternativ spleising kan DNA-oversetting hos mange dyr gjennomgå RNA-redigering -her, som kan endre aminosyre-sekvensen, og dermed funksjon til resulterende protein. F.eks. er det i blekkspruter funnet omfattende RNA-redigering for å tilpasse seg omgivelser, f.eks. kulde -her (41). Nylige studier har også avdekket omfattende RNA-redigering i mennesker. (42) For proteiner gjelder at selv om deres funksjon avhenger av deres tre-dimensjonale form, så bestemmes ikke alltid formen av aminosyre-sekvensen. Noen proteiner tilpasser lignende foldede former, til tross for meget ulike aminosyre-sekvenser. (43) Andre antar ulike former, til tross for at de har samme eller meget like aminosyre-sekvenser. Det siste er kjent som 'forvandlings' (metamorphic) proteiner. (44)
Endelige endres de fleste plante og dyre-proteiner kjemisk ved tilføyelse av sukker-molekyler, avhengig av organismens behov. Som ulike bio-kjemikere har uttrykt det, er resultatet et kompleksitetsnivå "som når utover genomet" (45) og "forsyner et tilleggs-nivå av informasjon i biologiske systemer." (46) Så påstanden "DNA spesifiserer RNA, som spesifiserer protein" er feilaktig. Men det er i siste steg av sentral-dogmet, ved å spesifisere oss, at det feiler mest dramatisk.
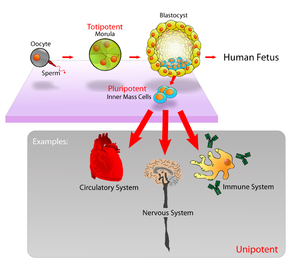
Bilde 5. Utvikling av ulike celletyper i menneskekroppen
Behovet for 3-dimensjonal informasjon
I de fleste tilfeller, etter at mRNA og proteiner er dannet, må de transporteres til spesifikke lokasjoner i celle, for at de skal fungere ordentlig. I tillegg til sin protein-kodende region, har noen mRNA en sekvens som kalles 'zip-kode', som spesifiserer adressen til hvor i cellen den skal transporteres. Men en RNA zip-kode, er lik et postnummer meningsløst, om det ikke tilsvarer en pre-eksisterende adresse. Celler med kjerne, inneholder mikroskopiske fibre (mikrotubuler), der molekylære motorer ferdes og transporterer ulik last gjennom cellen. Noen destinasjoner spesifiseres ved proteiner i celle-membranen. Selv om molekyler i celle-membranen er spesifisert av DNA, så er ikke 3D-arrangeringen av dem det. Biologiske membraner innehar essensiell romlig (3D-) informasjon, som ikke kan reduseres til informasjonen i DNA-sekvenser. (50)
I følge den britiske biologen Thomas Cavalier-Smith, er ideen at genomet inneholder all nødvendig informasjon for å danne en organisme, "rett og slett feil". Membran-mønstre spiller en nøkkelrolle i mekanismene som overfører lineær DNA-informasjon over til tre-dimensjonal former for enkeltceller og multicellulære organismer. Utvikling av dyr danner en kompleks, tredimensjonal multicellulær organisme, ikke ut fra lineær informasjon i DNA, men alltid ut fra en allerede kompleks, tredimensjonal encellet-organisme, det befruktede egget." (51)
 En slik kritikk får støtte også fra den kanadiske embryologen Brian C. Godwin, som forsøkte å forstå formdannelse (morfogenese). I 1985 skrev han at å trekke fram et genetisk DNA-program feiler i å adressere hovedproblemet, nemlig hvordan spesifikk romlig orden inntreffer i embryoer." (52) I 1996 skrev han: "Det er klart at spesifisering av form krever mer enn spesifisering av geno-type, noe som falsifiserer genetisk determinisme." (53) I 1999 publiserte den italienske genetikeren Giuseppe Sermonti boka 'Dimenticare Darwin', som ble oversatt til 'Why is a Fly not a Horse?' (2005) I boka utfordret Sermonti sentral-dogmet, og spurte hvorledes det hadde seg at helt annerledes dyr hadde lignende utviklings (developmental) gener, hvorfor er de da så annerledes?
En slik kritikk får støtte også fra den kanadiske embryologen Brian C. Godwin, som forsøkte å forstå formdannelse (morfogenese). I 1985 skrev han at å trekke fram et genetisk DNA-program feiler i å adressere hovedproblemet, nemlig hvordan spesifikk romlig orden inntreffer i embryoer." (52) I 1996 skrev han: "Det er klart at spesifisering av form krever mer enn spesifisering av geno-type, noe som falsifiserer genetisk determinisme." (53) I 1999 publiserte den italienske genetikeren Giuseppe Sermonti boka 'Dimenticare Darwin', som ble oversatt til 'Why is a Fly not a Horse?' (2005) I boka utfordret Sermonti sentral-dogmet, og spurte hvorledes det hadde seg at helt annerledes dyr hadde lignende utviklings (developmental) gener, hvorfor er de da så annerledes?
I 2003 påpekte cellebiolog og filosof Lenny Moss at 'biologisk orden er distribuert over flere paralelle og gjensidig avhengige systemer, slik at verken noe system, enn si molekyl, kunne tilordnes status som program/blueprint, sett av instruksjoner etc. for resten." (55) Om gener er ombyttbare over ordener og slekter, .., så må visselig spesifisiteten ved organismer bestemmes på et høyere nivås organisering." (56) I 2000 skrev MIT professor Evelyn Fox "I omtrent 50 år har vi innbilt oss at ved å oppdage molekylær basis for genetisk informasjon, så hadde vi funnet livets hemmelighet." Men "å benytte genet som kjerne-forklaring på biologisk struktur og funksjon, er mer ett trekk ved det 20. århundre, enn det vil bli i det 21."
Bilde 6. Hox-gen i banan-flue
Grunnen til at det er slik, argumenterte hun, er at programmet for levende organismer "består av, og lever i, et interaktivt kompleks, laget av genomiske strukturer og et svært nettverk av cellulært maskineri, som disse strukturene befinner seg i. Det kan endog være at dette programmet er ikke-reduserbart, ved at ikke noe mindre enn organismen selv, er i stand til å utføre jobben." (58) I så fall er det umulig at det kan ha utviklet seg til nåværende stadium, ett mutasjonstrinn om gangen, genetisk eller på annet vis? Zombi-vitenskap lunter forbi dette spørsmålet uten å ta det seriøst. Mutasjoner i bananfluer har lært oss noe om utviklings-genetikk, men ikke noe om hvordan evolusjon kan bygge ny form og funksjon. Uansett hva vi gjør med DNA i ei bananflue, er det 3 muligheter: ei normal bananflue, ei defekt bananflue eller ei død bananflue. Ikke en hestebrems, mye mindre en hest.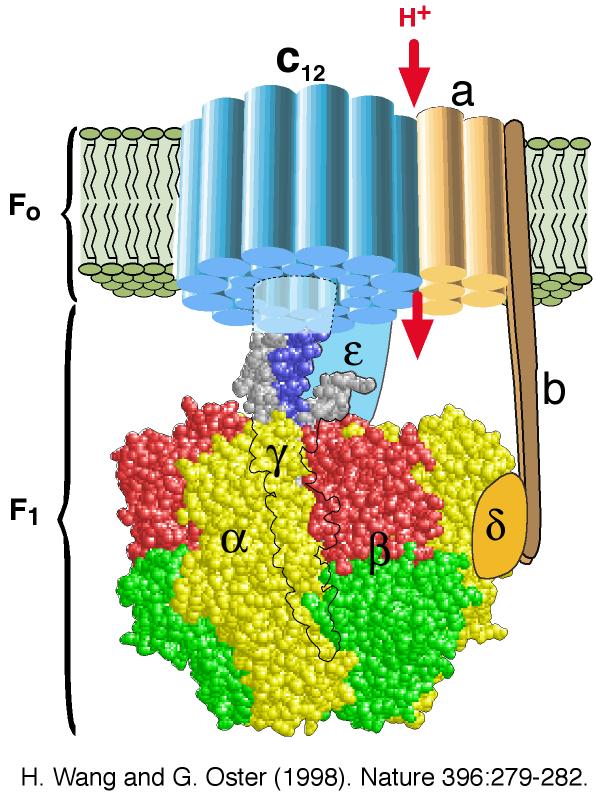
I en artikkel fra 2008, skrev Noble: "Det gir ikke fornuft i at genet opererer uavhengig av det øvrige cellulære maskineriet. Så, hvis et slikt nettverk er et program, så er det ikke et DNA-program. (60) Den amerikanske celle-biologen Stuart Newman kritiserte ideen at DNA er livets hemmelighet (2013): "Grunnen til at gener ble forhøyet til denne status i 1.omgang, var basert på mekanistisk ideologi som allerede var for nedadgående i legevitenskapene, enn på noe bevis for deres skapende evner." Men nå er det "gen-deterministiske synet kommet i en skarp nedadgående kurve i de sentrale biologiske vitenskapene." (61) Dessverre har mange ikke 'fått med seg dette', verken i media, populær-kulturen eller endog enkelte vitenskapsfolk. De fungerer som ett ekko fra svunne tider, og selv om de erkjenner at helheten er mer enn summen av delene, så fortsetter det meste som før. Ikke overraskende 'synger' R. Dawkins på dette 'refrenget'. I følge ham er det ufortjent at "denne trendy tingen" epigenetikk i bred forstand, "får all oppmerksomhet og ikke fortjener det." (63)
Også de siste par år er det kommet forskningsartikler, der media hauser opp innholdet, fra at "to proteiner aktiverte blueprintet som trengs for utvikling av organismer," (67,68) til at "to nøkkel-protein identifisert, som fungerer som genetiske arkitekter ved å danne blueprintet som trengs for tidligste stadier av embryo-utvikling." En lineær, sekvensiell en-dimensjonal DNA-kode kan vanskelig stå for den 3-dimensjonale romlige koden som trengs for utvikling av befruktede egg. Vedvarende tro på dette har grunnlag i materialisme. Feilen ligger ikke i genene, men i en form for vitenskap som kun godtar naturlige årsaker, uansett bevis. Når mange vitenskapsfolk forutsetter at leter man lenge nok, vil man finne en naturalistisk forklaring, samme hva en undersøker. Men forutsetningen om at det finnes naturlige forklaringer for alt, er ikke bare et utsagn om metode. Det fører til filosofisk naturalisme, som erkjenner materielle objekter og fysiske krefter som det eneste virkelige. Da er man (i så fall) over i zombi-science.
Bilde 7. Én av cellens mange nanomolekylære motorer (ATP/energi-motor)
Tillegg fra samtale mellom J. Wells og J. West -herfra:
DNA viser ikke hele sannheten. Det er et område for undersøkelse: Et gen kan ha 5-6 ulike beskjeder. Det finnes kode innen kode innen kode, slik at en beskjed kan leses på 5-6 'ulike språk'. Innen sukker-kode kan tusener av RNA-typer kodes. Sukkerkoden kan sml. med et postnummer-kode. I tillegg har vi membran-kode, elektroniske-koder, elektriske felt omkring embryoet etc. Tusenvis av RNA-strenger kan kodes fra en DNA-streng.
En kan snakke om ID i klasserommet, men da 'må' det snakkes nedsettende om det. Studenter er nysgjerrige, og om noe går mot lærebøkene, så vil de kanskje begynne å stille spørsmål, og gjøre folk oppmerksomme på at moderne forskning ofte går i mot dette foreldede synet.
Kilder:
20. Fr. Jacob, The Logic of Life oversatt av Betty E. Spillmann (Princeton:Princeton University Press, 1973), 3
23. H. Curtis og N.Sue Barnes, Biology, 5.utg (New York: Oxford University Press, 1976), 2, 23, 25
24. E. Mayr, Animal Species and Evoulution (Cambridge MA: Harvard University Press, 1963), 263
25. E. Mayr, Population Species and Evolution: An Abridgment of Animal Species and Evolution (Cambridge: MA: Harvard University Press, 1970), 162
26. Sean B. Carroll, Endless Forms Most Beautiful: The New Science of EvoDevo (New York: W.W. Norton, 2005), 294
32.Hall, Evolutionary Development Biology, 150
36. S.W.Herring, "Formation of the vertebrate face: Epigenetic and functional influences," American Zoologist 33 (1993): 472-483.
37. Eva Jablonka and M.J.Lamb, "The changing concept of epigenetics,m" Annals of the New York Academy of Sciences 981 (2002): 82-96
39.Wei Sun, X. You et al. "Ultra-deep profiling of alternativly spliced Drosphila Dscam isoforms by circularization-assisted mult-segment sequencing," EMBO Journal 32 (2013): 2029-2038.
40. Y. Barash et al, "Deciphering the splicing code", Nature 465 (2019: 53-59
41. S. Garret and J.J.C Rosenthal, "RNA-editing underlies temperature adaption in K+ Channels from polar octopuses," Science 335 (2012): 848-851
42. Jae Hoon Bahn et al. "Genomic analysis of ADAR1 binding and its involvement in multiple RNA processing pathways," Nature Communicatios 9 (2015): 6355
43. R.B.Russell and G.J. Barton, "Structural features can be unconserved in proteins with simular folds," Journal of Molecular Biology 244 (1994): 332-350
44. A.G.Murzin, "Metamorphic proteins," Science 320 (2008):1725-1726
45.R.G.Spiro, "Protein glycosylation: Nature Distribution, enzymatic formation, and disease implications of glycopeptide bonds," Glycobiology 12 (2002): 43R-56R
46. K.W.oremen et al, "Vertebrate protein glycosylation: Diversity, syntethesis and function," Nature Review Molecular Cell Biology 13 (2012): 448-462
50. J. Wells, "Membran patterns carry ontogenetic information that is specified independently of DNA," Bio-Complexity 2 (2014): 1-28
51. Th. Cav-Smith, "The memranome and membrane heredity in development and evolution," In Organelles, Genomes and Eukaryote Phylogeny, CRC Press (2004), 335-351
52. Br. C. Goodwin "What are the causes of morphogenesis?" BioEssays 3 (1985):32-36
53. G. Webster and Br. Goodwin, Form and Transformation: Generative and Relational Principles in Biology (Cambridge, Cambridge University Press, 1996), 134
55. L.Moss, What Genes Can't Do (Cambridge: MA: MIT Press, 2003), xviii
56. L.Moss, What Genes Can't Do (Cambridge: MA: MIT Press, 2003), 95-96
58. D.B. Wake, "Homplasy, homology and the problem of 'sameness' in biology", in Novartis Symposium, 222 -Homology, red. G.K.Bock og G. Cardew UK: John Wiley & Sons, 1999), 24-46. Se s.45.
60. D. Noble, "Genes and Causation," Philosophical Transactions of the Royal Society of London A 266 (2008): 3001-3015
61. Stuart A. Newman, "The Demise of the gene," Capitalism Nature Socialism 24 (2013): 6272
63. J. Webb, "The gene's still selfish: Dawkins famous idea turns 40," BBC News http://www.bbc.com/news/science-environment-36358104
67. Liz Williams, "Scientists uncover gene architects responsible for bodys bluesprint," /alter?Eliza Hall Institute of Medical Research (april 14, 2015):
http://www.wehi.edu.au/news/scientists-uncover-gen-architechts-responsible-bodys-bluesprint
68. Bilal N. Sheikh et al. "MOZ and MBI1 play opposing roles during Hox gene activation in ES cells oand in body segment identity specification in vivo," Proceedings of the National Academy of Sciences USA 112 (2015): 5437-5442
Stoffutvalg og bilder (untatt det 1) ved Asbjørn E. Lund