 Regulering og balanse
Regulering og balanse
(utdrag og refleksjoner ut fra ch.10 i: 'The Epigenetics Revolution'')
Et gen er et område av DNA-tråden der basene A,T,C og G er satt sammen i en bestemt rekkefølge (se Bilde 1). Genene er ordnet på rekke og rad. Genene utgjør ca 1,2 prosent av DNA-trådene. 'Genom' brukes gjerne om en enkelt celles totale DNA-innhold; omfatter DNA i cellekjernen pluss den lille mengden DNA som er lokalisert til mitokondriene. Vi begynner å forstå at ikke bare rekkefølgen i DNA-koden er viktig. Hva betyr den store delen: 98-99 prosent av genomet- som ikke er gener? Av de resterende 98-99% DNA, vet vi nå at mye er ansvarlig for regulering, opprettholdelse og omprogrammering av genetiske sekvenser. I tillegg inneholder de meget mobile segmenter av DNAet, som har fått navnet transposoner, fordi de kan framstille kopier av seg selv, og deretter flytte rundt til ulike steder på genomet. Det kan slukke eller aktivere visse gener. Det samme gjelder i stor grad for lange ikke-protein-kodende (ipk) RNA, og som vi skal se, for micro-RNA. Dessuten benyttes ikke-protein-kodende (ipk) DNA til DNA-profiler i rettsmedisin. Her har hvert menneske en unik profil.
 Bilde 1. Kromosominnhold
Bilde 1. Kromosominnhold
Vi så i forrige innlegg (lenke) på at lange ipkRNA kunne ha uforutsette og betydelige effekter på hvordan gener uttrykkes. Det kan gjøre at noen tror det må 'svære greier' til for å kunne utrette dette. Lange ipkRNA har betydelig innflytelse på cellen, men det er en annen like viktig virkning fra korte (20-24 basers lengde) ipkRNAs, og de er siktet inn mot mRNA (ikke DNA). Det fant man først ut i C. elegans ormen (referert tidligere). En av nøkkel regulatorene i C. elegans, er ett protein kalt LIN-14. Det tilhørende LIN-14 genet er mye uttrykt, ved at mye LIN-14 protein produseres. I hvert fall i tidlig embryo-fase, men det blir nedregulert i det ormen går fra stadium 1 til 2 i larvefasen.
Bilde 2. Innvirkning på LIN-14
Hvis LIN-14 genet muteres, blir det feil timing på ulike stadier. Om LIN-14 proteinet er slått på for lenge, starter ormen å repetere tidligere stadier. Dersom LIN-14 proteinet mistes for tidlig, går ormen for tidlig inn i senere larvestadier. Uansett blir ormens vekst mye forstyrret, og normale voksne strukturer utvikler seg ikke. I 1993 viste to uavhengige lab-rapporter hvordan uttrykket av LIN-14 ble kontrollert (17,18). Uventet var nøkkel-begivenheten binding av et lite ipkRNA til LIN-14 mRNA-molekylet. Det viser eks. på stilning av gen etter transkripsjon (avskrift) av genet. mRNA dannes, men hindres i å generere protein. Det blir en annen måte å kontrollere gen-uttrykk på enn hva tilfellet var for lange ipkRNA.
Dette arbeidet åpnet forståelse for denne måten å regulere gen-uttrykk på. Små ipkRNA vites nå å være en mekanisme som benyttes over hele plante og dyreriket, for å kontrollere gen-uttrykk. Det er ulike typer av små ipkRNA, men vi skal i hovedsak betrakte microRNA (miRNA). Minst 1000 ulike miRNA er identifisert i pattedyrs celler. miRNAer omkring 21 baser lange. Og de fleste synes å virke slik at de stilner genet etter transkripsjon. I stedet for å stoppe produksjon av mRNA, regulerer de hvordan mRNA opptrer. De gjør dette typisk ved å binde seg til den 3' 'ikke-oversatte' regionen, -på slutten av mRNA-molekylet-se Figur 1-under. De er til stede i det modne mRNA, men koder ikke for noen aminosyrer.
Figur 1. Regulering av utvikling (Lin 14)
 Ved spleising blir introner fjernet for å danne et mRNA som koder for protein. Men på begynnelsen (5' UTR) og slutten (3' UTR) av RNA-et, er det RNA som ikke koder for aminosyrer. Disse spleises heller ikke ut som introner. I stedet blir disse ikke-kodende regioner beholdt på det ferdige mRNA, og virker som regulerende sekvenser. Én av funksjonene til 3' UTR er å knytte seg til regulerende molekyler, inkludert miRNA. Disse samhandler bare om de 'gjenkjenner' hverandre, og da knytter de seg til hverandre på normalt vis: G med C, og A med U (tilsvarende T i DNA). Men nøkkel-regionen som de knytter seg til hverandre, er posisjon 2 til 8 på miRNA, de trenger ikke å matche mRNA over alle de 21 nukleotidene. Selv om matchen ikke trenger være varig, hindres oversettelse av mRNA til protein, så lenge den varer. Om matchen derimot er perfekt, så trigger binding av miRNA til mRNA ødeleggelse av mRNA ved enzymer som knytter seg til miRNA (19).
Ved spleising blir introner fjernet for å danne et mRNA som koder for protein. Men på begynnelsen (5' UTR) og slutten (3' UTR) av RNA-et, er det RNA som ikke koder for aminosyrer. Disse spleises heller ikke ut som introner. I stedet blir disse ikke-kodende regioner beholdt på det ferdige mRNA, og virker som regulerende sekvenser. Én av funksjonene til 3' UTR er å knytte seg til regulerende molekyler, inkludert miRNA. Disse samhandler bare om de 'gjenkjenner' hverandre, og da knytter de seg til hverandre på normalt vis: G med C, og A med U (tilsvarende T i DNA). Men nøkkel-regionen som de knytter seg til hverandre, er posisjon 2 til 8 på miRNA, de trenger ikke å matche mRNA over alle de 21 nukleotidene. Selv om matchen ikke trenger være varig, hindres oversettelse av mRNA til protein, så lenge den varer. Om matchen derimot er perfekt, så trigger binding av miRNA til mRNA ødeleggelse av mRNA ved enzymer som knytter seg til miRNA (19).
Bilde 3. Alternative spleiser fra pre-mRNA
Ett enkelt miRNA kan regulere mer enn ett mRNA. Vi så tidligere at ett gen kunne kode for masse ulike protein molekyler, ved å endre måten mRNA ble spleiset. Ett enkelt miRNA kan influere mange av disse spleisede versjonene samtidig. Alternativt kan ett miRNA også influere ganske urelaterte proteiner, kodet for av ulike gener, men som har like 3' UTR-sekvenser. Dette gjør det vanskelig å avdekke hva miRNA gjør i cellen, og effektene varierer i henhold til celle-type og alle andre gener (både protein-kodende, og ikke-protein-kodende), som cellen uttrykker til enhver tid. Fordi miRNA kan regulere masse andre gener, så kan virkningene av å bryte opp miRNA-kopiering bli meget omfattende. Dette har også viktige helsemessige implikasjoner.
 Det faktum at det menneskelige genom består av 98% ikke-protein-kodende DNA, kan antyde at det er et svært potensiale her. Noen forfattere har antydet at ipkRNA er nøkkelfaktoren for å utvikle menneskets største skillende trekk, våre høyere tanke-prosesser (20). I 2005 ble det utgitt en gjennomgang av genomet for sjimpanse. Statistikk influeres av hvorvidt repeterende regioner forekommer én eller flere ganger. {Den påvirkes også hvorvidt hvilke regioner en sammenligner, og hvor selektivt en går til verks -lenke; oversetters merknad.} Selv om genomene kan synes like, er det kun 1/3 del av proteinene som er eksakt like, mens resten kan skille seg med noen få aminosyrer. Som nevnt tidligere har vi også til felles at 98% av genomet ikke koder for proteiner. Dette antyder at begge arter benytter ipkRNA til å danne komplekse, regulerende nettverk for å styre uttrykket av gener og proteiner. Det er funnet én spesifikk forskjell i hvordan ipkRNA blir behandlet i cellene til de to artene.
Det faktum at det menneskelige genom består av 98% ikke-protein-kodende DNA, kan antyde at det er et svært potensiale her. Noen forfattere har antydet at ipkRNA er nøkkelfaktoren for å utvikle menneskets største skillende trekk, våre høyere tanke-prosesser (20). I 2005 ble det utgitt en gjennomgang av genomet for sjimpanse. Statistikk influeres av hvorvidt repeterende regioner forekommer én eller flere ganger. {Den påvirkes også hvorvidt hvilke regioner en sammenligner, og hvor selektivt en går til verks -lenke; oversetters merknad.} Selv om genomene kan synes like, er det kun 1/3 del av proteinene som er eksakt like, mens resten kan skille seg med noen få aminosyrer. Som nevnt tidligere har vi også til felles at 98% av genomet ikke koder for proteiner. Dette antyder at begge arter benytter ipkRNA til å danne komplekse, regulerende nettverk for å styre uttrykket av gener og proteiner. Det er funnet én spesifikk forskjell i hvordan ipkRNA blir behandlet i cellene til de to artene.
Det har å gjøre med editering av ipkRNA (22). Menneskelige celler synes å ha vanskelig for å unnlate å modifisere det videre ved ulike mekanismer. Helt spesielt vil menneskelige celler ofte endre basen A til en som kalles I, for Inosin. Base A binder seg som kjent til T i DNA, eller U i RNA. Men base I kan også danne par med C eller G! Dette endrer klart sekvensene som en ipkRNA kan knytte seg til og dermed regulere. Vi mennesker kan editere våre ipkRNA-molekyler på en måte som ingen pattedyr kan (23). Spesielt skjer dette i hjernen. Om en skal se etter faktorer som påvirker hvordan mennesker er så mye mer 'mindful' enn sjimpanser, selv om vi har mye DNA-maler felles.
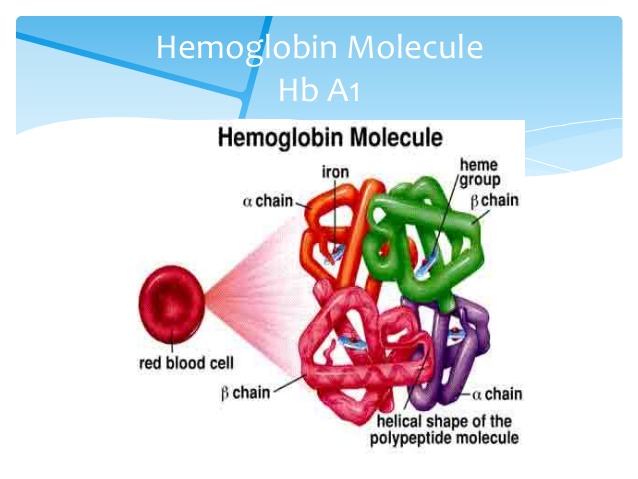
Det er rasjonelt å benytte ipkRNA til slikt, da våre proteiner er delikate mekanismer, som tåler lite slingringsmonn. F.eks. har hemoglobin som transporterer de røde blodcellers oksygen rundt i kroppen, aldri blitt utkonkurrert av noe funnet opp i et laboratorium. Om hemoglobin-molekyl ødelegges i mutasjoner, kan det føre til sigdcelle-anemi. Det samme er tilfellet for de fleste proteiner. De fleste proteiner er så gode som de kommer til å bli. Ved å endre regulering av proteiner, kan en påvirke når og i hvor stor grad spesifikke proteiner blir uttrykt. Det foreligger dokumentasjon på at dette faktisk finner sted.
 Et syndrom i mennesker, DiGeorge, innebærer at ca. 3 millioner baser har gått tapt fra én av kopiene i kromosom 22 (40). Denne regionen inneholder mer enn 25 gener. Ett av genene er kalt DGCR8, og tilhørende protein er essensielt for normal produksjon av miRNA. Genetisk modifisert mus, med bare en funksjonell kopi av DGCR8, utvikler kognitive problemer, spesielt i læring og rom-avstands prosesser (41). Det understøtter idéen at miRNA kan være viktige i nevrologiske prosesser. Vi vet at ipkRNA er viktige i kontroll av cellulært mangfold (pluripotency) og differensiering. Det er et lite sprang til en hypotese om at miRNA kan være viktige i utvikling av kreft. Kreft er typisk en sykdom hvor cellene holder på å formere seg, som har paralleller hos stamceller. Kreftsvulster ser også nokså udifferensierte og uorganiserte, i motsetning til fullt organiserte og velorganisert framtreden av normalt, sunt cellevev. Det er nå et sterk bevismengde for at ipkRNA spiller en rolle i kreft. Det kan innebære enten tap av utvalgte miRNA eller over-produksjon av andre miRNA.
Et syndrom i mennesker, DiGeorge, innebærer at ca. 3 millioner baser har gått tapt fra én av kopiene i kromosom 22 (40). Denne regionen inneholder mer enn 25 gener. Ett av genene er kalt DGCR8, og tilhørende protein er essensielt for normal produksjon av miRNA. Genetisk modifisert mus, med bare en funksjonell kopi av DGCR8, utvikler kognitive problemer, spesielt i læring og rom-avstands prosesser (41). Det understøtter idéen at miRNA kan være viktige i nevrologiske prosesser. Vi vet at ipkRNA er viktige i kontroll av cellulært mangfold (pluripotency) og differensiering. Det er et lite sprang til en hypotese om at miRNA kan være viktige i utvikling av kreft. Kreft er typisk en sykdom hvor cellene holder på å formere seg, som har paralleller hos stamceller. Kreftsvulster ser også nokså udifferensierte og uorganiserte, i motsetning til fullt organiserte og velorganisert framtreden av normalt, sunt cellevev. Det er nå et sterk bevismengde for at ipkRNA spiller en rolle i kreft. Det kan innebære enten tap av utvalgte miRNA eller over-produksjon av andre miRNA.
Bilde 5. Redusere overrepresentasjon eller øke underrepresentasjon -i forb. med miRNA
Kreft er en sykdom som utvikles i flere trinn, der flere må gå feil før en kommer til kreft. Kronisk lymfekreft er den vanligste kreftform i mennesker. Ca. 70% av tilfellene av denne sykdommen, har mistet ipkRNA (miR-15a og miR-16-I). At såpass mange tilfeller manglet disse spesifikke miRNA antyder at tap av disse sekvensene inntraff tidlig i utvikling av sykdommen. Et eks. på den alternative mekanismen, overrepresentasjon av miRNA i kreft, er tilfellet med miR-17-92 opphopningen. Denne opphopningen er over-uttrykt i en rekke kreftformer (43). Det er et betydelig antall rapporter som nå er publiser om anormale uttrykk av miRNA i kreft (44). I tillegg, er det et mutert gen (TARBP2), i noen nedarvede kreftformer (45). TARB2 proteinet er involvert i normal foredling av miRNA. Det styrker ytterligere rollen for miRNA i initiering og utvikling av visse kreftformer.
 Siden en stadig større datamengde antyder en hovedrolle for miRNA i kreft, så er det ikke overraskende at vitenskapsfolk begynner å bli opptatt av muligheten for å benytte disse molekylene til å behandle kreft. Idéen ville være å erstatte manglende miRNA, eller å hindre noen som var overrepresentert. Håpet var at dette kunne oppnås ved å dosere kunstige varianter av miRNAs til kreftpasienter med ubalanse av miRNAs. Det kunne også ha effekt i andre sykdommer, der miRNAs var i ubalanse. Det finnes også en annen variant siRNAs (Small Interfering RNAs), som benytter samme prosesser som miRNAs for å undertrykke gener. Dette har vært benyttet av store medisinske firma, med varierende resultater. Ett av problemene har vært å skape medisiner ut fra nukleinsyre, da et gjennomsnittlig siRNA-molekyl er over 50 ganger større enn f.eks. ett ibuprofen-molekyl. Desto større et medisinpreparat er, desto større problemer er det gjerne i å få store nok doser i pasienten, og å beholde dem lenge nok til å virke. Så har en også begynt med andre typer behandlinger, der kroppens egne antistoffer spiller en sentral rolle i å bekjempe kreft, selv om disse er begrenset fordi antistoffene bare kan gjenkjenne proteiner på celleoverflaten.
Siden en stadig større datamengde antyder en hovedrolle for miRNA i kreft, så er det ikke overraskende at vitenskapsfolk begynner å bli opptatt av muligheten for å benytte disse molekylene til å behandle kreft. Idéen ville være å erstatte manglende miRNA, eller å hindre noen som var overrepresentert. Håpet var at dette kunne oppnås ved å dosere kunstige varianter av miRNAs til kreftpasienter med ubalanse av miRNAs. Det kunne også ha effekt i andre sykdommer, der miRNAs var i ubalanse. Det finnes også en annen variant siRNAs (Small Interfering RNAs), som benytter samme prosesser som miRNAs for å undertrykke gener. Dette har vært benyttet av store medisinske firma, med varierende resultater. Ett av problemene har vært å skape medisiner ut fra nukleinsyre, da et gjennomsnittlig siRNA-molekyl er over 50 ganger større enn f.eks. ett ibuprofen-molekyl. Desto større et medisinpreparat er, desto større problemer er det gjerne i å få store nok doser i pasienten, og å beholde dem lenge nok til å virke. Så har en også begynt med andre typer behandlinger, der kroppens egne antistoffer spiller en sentral rolle i å bekjempe kreft, selv om disse er begrenset fordi antistoffene bare kan gjenkjenne proteiner på celleoverflaten.
Bilde 6. miRNA kan også benyttes til å spore/identifisere kreft
Referanser:
17. Lee et al. (1993), cell 75: 843-854
18. Wightman et al. (1993), Cell 75: 858-862
 19. Bartel (2009, Cell 136: 215-233
19. Bartel (2009, Cell 136: 215-233
20. Mattick, J.S. (2010), BioEssays 32: 548-552
21. Chimpanzee Sequencing and Analysis Concortium (205); Nature 437: 69-87
22. Athanasiasdis et al. (2004), PLoS Biol. 2: e91
23. Paz-Yaacom et al. (2010)), Proc Natl Acad Sii. USA 107: 12174-9
40. DiGeorge syndrom: http://omim.org/entry/188400
41. Strax et al. (2008) Nature Genetics 38: 813-818
43. Volinia et al. (2006), Proc Natul Acad Sci. USA 103: 2257-2261
44. Garzon et al. (2010), Nature Reviews Drug Discovery 9: 775-789
45. Melo et al. (2009), Nature Genetics 41: 365-370
Bilde 7. Kreft-utvikling og motstand
Stoffutvalg og bilder ved Asbjørn E. Lund