Komplekse tilpasninger og neo-darwinistisk matematikk -fortsetter
(Fritt utvalg fra Darwins Doubt; S.C.Meyer, Harper One, 2013, Ch.12B) {med oversetters kommentar i klamme-parentes}
Så mange endringer på liten tid-repetisjon
 Michael Behe og David Snoke hadde benyttet prinsippene fra populasjonsgenetikk, for å fastsette sannsynligheten til ulike antall koordinerte (samordnede) mutasjoner i en gitt tidsperiode. De anvendte standard neo-darwinistisk modell, med gen-duplisering og påfølgende mutasjoner. De spurte: "Er det mulig at det er nok tid i evolusjonær historie til å generere koordinerte (samordnede) mutasjoner? Om så var, hvor mange koordinerte mutasjoner er det sannsynlig å forvente i en gitt tidsperiode, gitt ulike populasjons-størrelser, mutasjonsrater og generasjonsvarighet? Så for ulike kombinasjoner av disse variable beregnet de hvor lang tid det ville ta å frambringe tre eller mer koordinerte (samordnede) mutasjoner. Det de fant ut generelt, var at sannsynligheten for multiple mutasjoner, som oppsto i nære funksjonelt relevante, koordinasjoner, var "avskrekkende" (prohibitively) liten. Det ville sannsynligvis ta ekstremt lang tid, typisk mye lenger enn alderen på jorda.
Michael Behe og David Snoke hadde benyttet prinsippene fra populasjonsgenetikk, for å fastsette sannsynligheten til ulike antall koordinerte (samordnede) mutasjoner i en gitt tidsperiode. De anvendte standard neo-darwinistisk modell, med gen-duplisering og påfølgende mutasjoner. De spurte: "Er det mulig at det er nok tid i evolusjonær historie til å generere koordinerte (samordnede) mutasjoner? Om så var, hvor mange koordinerte mutasjoner er det sannsynlig å forvente i en gitt tidsperiode, gitt ulike populasjons-størrelser, mutasjonsrater og generasjonsvarighet? Så for ulike kombinasjoner av disse variable beregnet de hvor lang tid det ville ta å frambringe tre eller mer koordinerte (samordnede) mutasjoner. Det de fant ut generelt, var at sannsynligheten for multiple mutasjoner, som oppsto i nære funksjonelt relevante, koordinasjoner, var "avskrekkende" (prohibitively) liten. Det ville sannsynligvis ta ekstremt lang tid, typisk mye lenger enn alderen på jorda.
 Bilde 1. Forutsetning for Behes og Snokes undersøkelse
Bilde 1. Forutsetning for Behes og Snokes undersøkelse
Det de argumenterte for i sin artikkel fra 2004, var at det å danne ett nytt protein, ofte krever mange usannsynlige mutasjoner samtidig. De tok i betraktning det usannsynlige i at flere, funksjonelt nødvendige mutasjoner fant sted samtidig. De fant at om å danne ett nytt gen krevde flere, koordinerte (samordnede) mutasjoner, så ville ventetiden øke eksponentielt for hver nødvendig mutasjons-endring. De anslo også, {ut fra populasjons-genetiske modeller} hvordan populasjonsstørrelser påvirket hvor lang tid det ville ta, å danne nye gener, dersom det krevde flere koordinerte mutajoner. Som ventet fant de ut at økning i populasjonsstørrelser, førte til reduksjon i fikseringstid og v.v. Et viktig funn var at selv om dannelse av ett nytt gen krevde 'bare' to koordinerte (samordnede) mutasjoner, så ville neo-darwinistiske mekanismer enten kreve enormt store populasjoner, og/eller ekstremt lange ventetider. Dersom koordinerte mutasjoner var nødvendige, så møtte evolusjon på genetisk nivå et 'catch-22' dilemma: enten trengtes urealistisk lange (ikke-mulig i jordisk sammenheng), eller enormt store populasjoner (på virusnivå i jordisk sammenheng). De uttrykte det slik: "tallene synes uansett uoppnåelige" (26)
Fig. 1. Sammenheng mellom antall koordinerte mutasjoner, populasjonsstørrelse og antall generasjoner som trengs for at de skal trenge gjennom i populasjonen.
Forklaring til Fig.1: diagrammet viser populasjons-størrelser og tid (målt i antall generasjoner), som er nødvendig for å produsere ett gen eller en egenskap, om dannelsen av dette krever flere koordinerte mutasjoner. Bemerk at enhver multi-mutasjons begivenhet, som krever flere enn to koordinerte mutasjoner, kan ikke utvikles ved gen-duplisering og påfølgende mutasjoner i en populasjonsstørrelse som har eksistert på jorda. For de fleste populasjoner, av normal (jordisk) størrelse, så er også dannelse av to koordinerte mutasjoner utenfor rekkevidde for neo-darwinistiske mekanismer.
Riktignok fant de at mutasjoner og seleksjon kunne danne to koordinerte mutasjoner på 'bare' 1 million generasjoner, men det var under forutsetning av en populasjonsstørrelse på 1 billion (1 million millioner) eller flere mutlicellulære organismer. Det er et antall som overstiger størrelsen på alle dyreslag som har levd på jorda til en hvilken som helst tid (27). Motsatt fant de at to koordinerte mutasjoner kunne forekomme i en populasjon på bare 1 million organismer, men bare om mekanismen hadde 10 milliarder generasjoner til sin disposisjon. Forutsatt at hver multicellulær organisme bare levde 1 år, ville det likevel kreve 10 milliarder år, -mer enn det dobbelte av jordas alder. Det er en ufornuftig tidsperiode å vente på att ett nyt gen skal dukke opp, for ikke å snakke om mer betydelige evolusjonære innovasjoner.
De fant imidlertid ett lite 'tilfluktssted' (skravert på figur til venstre): Ett slik gen kunne formodentlig dannes i en populasjon på 1 milliard organismer, på 'bare' 100 millioner generasjoner. Men om populasjonen som utvikler seg gjennom to-mutasjons-trekk, har færre enn 1 milliard organismer, så øker ventetiden urealistisk lenge. Uansett gjelder disse tallene bare det tilfellet der bare to koordinerte mutasjoner er nødvendige for å bygge et nytt gen. Dersom det krevde flere enn to koordinerte mutasjoner, så var urealistisk lange ventetider nødvendig, for alle realistiske populasjonsstørrelser. For tre eller flere koordinerte mutasjoner, var det ingen 'tilfluktssteder' tilgjengelig. Sålede konkluderte de at mekanismen med gen-duplisering og punktmutasjoner alene, ville være ineffektiv, i det minste for multi-cellulære organismer. (28)
Behe og Snoke fant altså, ved å anvende prinsippene anvendt i populasjons-genetikk at den standard-evolusjonistiske modellen for gen-evolusjon, ikke strakk til som foklaring, der det krevdes flere enn to koordinerte mutasjoner, i multicellulære, eukaryote organismer.
 I 2007 utga Michael Behe boka 'The Edge of Evolution', der han utdypet resultatene fra artikkelen i 2004. I det han brukte offentlige helse-data, knyttet til resistens mot malaria-medisinen qloroquine, i den encellede organismen som forårsaker malaria. Behe ga på det viset en annen innfallsvinkel til støtte for konklusjonen om at multiple, koordinerte mutasjoner ofte er nødvendige for å produsere selv mindre genetiske tilpasninger. Behe sluttet ut fra disse offentlige helse-dataene at resistens mot qloroquine inntreffer bare i én av (10 opphøyd i 20) malaria-fremkallende celler. Ut fra dette resultatet sluttet han bakover at resistensen krevde flere, om ikke nødvendigvis, koordinerte (samordnede) mutasjoner for å framkalles. Han kalte dette trekket et 'qloroquine complexity cluster', eller ett CCC (29). Behe ønsket å utforske 'evolusjonens grense', grensen for den skapende kraft til utasjoner og seleksjon på genetisk nivå. I det han hadde funnet at denne grensen kunne nåes på relativt kort tidsrom, så lurte han på hvor lenge det ville ta å danne trekk med større kompleksitet i ulike populasjonsstørrelser.
I 2007 utga Michael Behe boka 'The Edge of Evolution', der han utdypet resultatene fra artikkelen i 2004. I det han brukte offentlige helse-data, knyttet til resistens mot malaria-medisinen qloroquine, i den encellede organismen som forårsaker malaria. Behe ga på det viset en annen innfallsvinkel til støtte for konklusjonen om at multiple, koordinerte mutasjoner ofte er nødvendige for å produsere selv mindre genetiske tilpasninger. Behe sluttet ut fra disse offentlige helse-dataene at resistens mot qloroquine inntreffer bare i én av (10 opphøyd i 20) malaria-fremkallende celler. Ut fra dette resultatet sluttet han bakover at resistensen krevde flere, om ikke nødvendigvis, koordinerte (samordnede) mutasjoner for å framkalles. Han kalte dette trekket et 'qloroquine complexity cluster', eller ett CCC (29). Behe ønsket å utforske 'evolusjonens grense', grensen for den skapende kraft til utasjoner og seleksjon på genetisk nivå. I det han hadde funnet at denne grensen kunne nåes på relativt kort tidsrom, så lurte han på hvor lenge det ville ta å danne trekk med større kompleksitet i ulike populasjonsstørrelser.
Bilde 2. 'Evolusjonens grense'
Han ba leserne å vurdere ett hypotetisk gen, dobbelt så komplekst som CCC. Det var en begivenhet som krevde to koordinerte hendelser, hver av dem så kompleks som CCC. Behe undret hvor lenge det ville ta å oppnå to slike hendelser, dersom begge hendelsene måtte inntreffe samtidig inne i den samme organismen, på koordinert (samordnet) vis, for å produsere den nye egenskapen. Ved å benytte prinsippene i populasjons-genetikk, viste han at det kom til å ta enormt mye lenger tid og/eller mange flere organismer, enn det som var rimelig i livets historie på jorda. Som eks. påpekte han at om (10 opphøyd i 20) organismer behøvdes for å oppnå ett tilfelle av CCC, så ville kvadratet av det antallet (10 opphøyd i 40) være nødvendig for at to koordinerte CCC-er skulle inntreffe, før det ga noen seleksjonsfordel (30). Nå er det bare det, at til sammen har det ikke levd flere enn (10 opphøyd i 40) organismer på jorda. Det innebærer at hele jordas samlede livshistorie, knapt ville være tilstrekkelig for at en slik begivenhet skulle inntreffe. (31) {Ikke rart at evolusjonister nå forsøker seg med at livet er medbragt fra meteoritter, bare synd at de 'glemmer' å opplyse at aktuelle aminosyrer kan dannes på naturlig vis -oversetters tilføyelse}.
Behe viste at problemet i forhold til koordinerte (samordnede) mutasjoner var spesielt stort for lenge-levende organismer, med relativt små populasjons-størrelser. F.eks. organismer som pattedyr, eller mer spesifikt mennesket og deres formodede før-menneskelige forfedre. Basert på relevante estimerte mutasjons-rater, kjente populasjonsstørrelser og generasjons-lengde, beregnet han tiden det ville ta for to koordinerte (samrodnede) mutasjoner, i menneskets slektslinje. Han kalkulerte at det ville ta flere hundre millioner år, bare å gjennomføre en slik begrenset endring. Likevel er mennesker og sjimpanser tenkt å ha skilt lag bare for ca. 6 millioner år siden. Behe sluttet dermed at neo-darwinistiske mekanismer ikke har tilgjengelig kapasitet til å forklare to koordinerte mutasjoner, på tiden tilgjengelig for menneskelig evolusjon. Dermed kan den heller ikke forklare hvordan mennesket oppsto. {Ikke rart at evolusjonister da har 'forskjøvet' stamfaren bakover på ubestemt tid, som en felles stamfar til aper og mennesker - -oversetters tilføyelse}.
Naturlig nok ville noen etterprøve Behes resultater: Snart etter publikasjonen av 'The Edge of Evolution', forsøkte to evolusjonistiske, matematiske biologer, Rick Durret og Deena Schmidt, å bestride Behes konklusjoner, ved å gjøre egne beregniner. Deres artikkel ("Waiting for Two Mutations: With Applications to Regulatory Sequence Evolution and the Limits of Darwinian Evolution," som også anvendte en modell basert på populasjons-genetikk for å kalkulere nødvendig tidsrom for å danne to samordnede (koordinerte) mutasjoner i menneskets livstre. Selv om de beregnet en kortere ventetid enn Behe, understreket likevel deres resultat det usannsynlige i å stole på neo-darwinistiske mekanismer, for å danne koordinerte mutasjoner i evolusjonær tidsskala.
I følge deres kalkulasjoner ville det ta 216 millioner år å generere to koordinerte mutasjoner i hominid-livslinjen. Men dette er 30 ganger mer tid enn aktuelt tidsrom for å danne mennesker og sjimpanser og alle deres distinkte, komplekse tilpasninger og ulikheter, fra den formodede 'felles stamfaren'. I den hensikt å gjendrive Behes påstander, førte deres arbeid til at hoved-resultatet ble bekreftet. Som de to, Durret og Deena Schmidt, påpekte så viste deres beregninger at det er "meget urealistisk at to eller flere koordinerte mutasjoner inntreffer i rimelig tidsrom" (32). Så kalkulasjoner fra både kritikere og tilhengere av neo-darwinismen viser nå, at dersom det er nødvendig med koordinerte mutasjoner for å danne nye gener og proteiner, så viser neo-darwinistisk matematikk selv, via prinsippene i populasjons-genetikken, at de neo-darwinistiske mekanismene er utilstrekkelige.
Uttesting av samvirke-alternativet
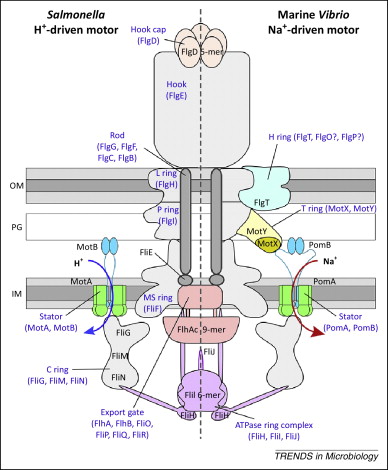 Men trenger det å generere nye gener og proteiner koordinerte mutasjoner? Behe og Snoke sluttet såpass, basert på et udiskutabelt faktum i molekylær biologi: Mange proteiner støtter seg til sett av aminosyrer, som handler i samordning (koordinasjon), for å utføre sine funksjoner. I tillegg argumenterte Behe i 'The Edge of Evolution' ut fra funksjonelt grunnlag, at mange biologiske systemer ville kreve koordinerte, tilpassede mutasjoner, siden fravær av bare ett eller noen få gen-produkter, ville få dem til å tape funksjonalitet. Behe viste spesifikt at atskillige molekylære maskiner inni celler, f.eks. bakterieflagellens motor (33), krever koordinert samhandling av flere protein-subsystemer for å vedlikeholde deres funksjon. Behe konkluderte likevel ikke med at nye gener og proteiner i seg selv, representerte komplekse tilpasninger.
Men trenger det å generere nye gener og proteiner koordinerte mutasjoner? Behe og Snoke sluttet såpass, basert på et udiskutabelt faktum i molekylær biologi: Mange proteiner støtter seg til sett av aminosyrer, som handler i samordning (koordinasjon), for å utføre sine funksjoner. I tillegg argumenterte Behe i 'The Edge of Evolution' ut fra funksjonelt grunnlag, at mange biologiske systemer ville kreve koordinerte, tilpassede mutasjoner, siden fravær av bare ett eller noen få gen-produkter, ville få dem til å tape funksjonalitet. Behe viste spesifikt at atskillige molekylære maskiner inni celler, f.eks. bakterieflagellens motor (33), krever koordinert samhandling av flere protein-subsystemer for å vedlikeholde deres funksjon. Behe konkluderte likevel ikke med at nye gener og proteiner i seg selv, representerte komplekse tilpasninger.
Bilde 3. Bakterieflagellens motor krever koordinert samordning
Noen neo-darwinister har foreslått en modell for protein-dannelse, kjent som 'samhandling' (co-option). I denne modellen, er et protein som utfører en funksjon endret eller samvirket til å utføre en annen handling. Denne modellen forestiller seg nye trekk, som krever flere mutasjoner, oppstår i en steg-for-steg måte for å produsere ett protein. La oss si protein B, fra et protein som manglet denne egenskapen, la oss si protein A. Ut fra andre slike 'fordels-grunner' blir ikke proteinet selektert bort,. I det mutasjoner fortsetter å generere nye proteiner med litt andre funksjoner, så ville de ha skapt et protein nært nok i sekvens og struktur, til at bare noen få tilleggsendringer trengs for å endre fra protein A til B.
 Douglas Axe og molekylær biolog-kollega Ann Gauger, som arbeider ved biologisk institutt i Seattle, besluttet å utsette denne hypotesen for en kreativ, eksperimentell test (34). Ved å gjøre det, ville de kunne finne ut hvorvidt evolusjon av nye trekk, typisk krever flere samordnede mutasjoner, eller om trekket i stedet kunne oppstå ved samhandling. Axe og Gauger gjennomsøkte databaser for å finne proteiner som var så nær hverandre i sekvens og struktur som mulig, men like fullt utførte ulike funksjoner. De identifiserte to proteiner som møtte disse kriteriene. Ett av disse proteinene, -Kbl2 trengs for å bryte ned en aminosyre (threonine) og den andre -BioF2, trengs for å danne ett vitamin (biotin). Kbl2 og BioF2, er enzymer som benytter ulike katalytiske mekanismer for å øke ulike kjemiske reaksjoner i E-coli bakterien. Gauger og Axe innså at om de kunne transformere Kbl2 til ett protein som utførte samme funksjon som BioF2, med bare ett eller meget å koordinerte aminosyre-endringer, så kunne det, avhengig av antall mutasjoner, demonstrere at to proteinene som var nær nok hverandre i funksjon og sekvens, kunne få til en funksjonsendring i evolusjonær tid, slik samhandlings-talsmennene forfektet. Om det derimot var nødvendig med mange mutasjons-endringer, så kunne det etablere, at darwinistiske mekanismer ikke kunne avstedkomme det funksjonelle spranget fra A til B i evolusjonær tid. Det ville innebære at en endog større grad av likhet mellom proteiner måtte være til stede, for at samvirke-hypotesen skulle være troverdig.
Douglas Axe og molekylær biolog-kollega Ann Gauger, som arbeider ved biologisk institutt i Seattle, besluttet å utsette denne hypotesen for en kreativ, eksperimentell test (34). Ved å gjøre det, ville de kunne finne ut hvorvidt evolusjon av nye trekk, typisk krever flere samordnede mutasjoner, eller om trekket i stedet kunne oppstå ved samhandling. Axe og Gauger gjennomsøkte databaser for å finne proteiner som var så nær hverandre i sekvens og struktur som mulig, men like fullt utførte ulike funksjoner. De identifiserte to proteiner som møtte disse kriteriene. Ett av disse proteinene, -Kbl2 trengs for å bryte ned en aminosyre (threonine) og den andre -BioF2, trengs for å danne ett vitamin (biotin). Kbl2 og BioF2, er enzymer som benytter ulike katalytiske mekanismer for å øke ulike kjemiske reaksjoner i E-coli bakterien. Gauger og Axe innså at om de kunne transformere Kbl2 til ett protein som utførte samme funksjon som BioF2, med bare ett eller meget å koordinerte aminosyre-endringer, så kunne det, avhengig av antall mutasjoner, demonstrere at to proteinene som var nær nok hverandre i funksjon og sekvens, kunne få til en funksjonsendring i evolusjonær tid, slik samhandlings-talsmennene forfektet. Om det derimot var nødvendig med mange mutasjons-endringer, så kunne det etablere, at darwinistiske mekanismer ikke kunne avstedkomme det funksjonelle spranget fra A til B i evolusjonær tid. Det ville innebære at en endog større grad av likhet mellom proteiner måtte være til stede, for at samvirke-hypotesen skulle være troverdig.
Bilde 4. Kbl(2) og BioF(2)
Etter grundig å ha undersøkt strukturelle likheter innen en stor klasse av strukturelt lignende enzymer, så visste de at Kbl2 og BioF2 var så lik hverandre i sekvens og struktur, som det var mulig for to proteiner med ulik funksjonalitet. Om det viste det seg at for å konvertere det ene proteinet til det andre, måtte det til mange koordinerte mutasjoner, flere enn de som kunne forventes å ville skje i aktuell tidsperiode, så ville det ha ødeleggende virkning for standard-utgavene av evolusjonsteorien. Om proteiner måtte ligne hverandre enda mer enn Kbl(2) og BioF(2), for at samvirke kunne skje, så ville i praksis samvirke-hypotesen ikke spille noen særlig rolle. Det er ikke mange kjente hopp, som er så små som det.
Axe og Gauger identifiserte først de aminosyrene som var mest sannsynlig, om de muterte, ville danne en overgang fra Kbl2-funksjon til BioF2-funksjon. Så muterte de systematisk fra sted til sted, enkeltvis og i grupper. inkl. ulike aminosyre-kombinasjoner. Resultatene var entydige. De fant at de ikke kunne fremkalle funksjons-endringen som de søkte, verken med én eller et lite antall aminosyrer. Selv ikke om de muterte større antall aminosyrer samtidig, altså flere koordinerte mutasjoner enn det som troverdig kunne ha skjedd, i hele den evolusjonære historie.
Selv om deres forsøk på å endre Kbl2 til BioF2 feilet, så feilet ikke eksperimentet. Det tillot dem for første gang å etablere det faktum, at samvirke-hypotesen for protein-evolusjon manglet troverdighet. Det kreves rett og slett for mange koordinerte mutasjoner å endre funksjon fra ett protein til ett annet, selv med ekstremt like proteiner. Dermed ville tiden som slike endringer kunne forventes å inntreffe på, ikke være i samsvar med evolusjonær historie. Axe utviklet en foredlet populasjonsgenetisk modell, til å beregne ulike ventetider. Resultatene hans samsvarte i grove trekk med Behes og Snokes. Om Axe tok hensyn til 'sannsynlighets fitness-kostnad' til en organisme som bar på unødige gen-duplikater, som var nødvendig i teorien, så kom han til at forventet ventetid for 3 koordinerte mutasjoner overstiger jordas alder. Derfor bestemte han en øvre grense på to koordinerte mutasjoner for det som kunne forventes å inntreffe i ett duplikat gen, gjennom jordas historie. Dersom en så bort fra fitness-kostnaden, kalkulerte han en øvre grense på 6 koordinerte mutasjoner. I eksperimentet deres, klarte ikke Axe og Gauger å tilvirke en funksjonell endring med mer enn 6 koordinerte mutasjoner {de fant at det krevdes minst 7; lenke}. Så det er lite som tyder på, verken ut fra forsøk eller beregninger, at samvirke-alternativet er troverdig.
Hva det hele innebærer
 Ved å vise det usannsynlige ved samvirke modellen for protein-evolusjon, bekreftet Axe og Gauger at gener og proteiner i seg selv representerer komplekse tilpasninger. De er avhengige av koordinert interaksjon av flere underenheter, som må oppstå som gruppe for å bevirke noen funksjonell fordel. Det innebærer igjen at biologer ikke bare kan anta at mutasjoner greit vil skape nye gener og trekk, slik det lenge har vært gjort. Både de biologer som er nevnt her, og flere andre , har nylig vist at å generere det antall multippelt-koordinerte mutasjoner som trengs endog til å produsere ett nytt gen, eller få to nærliggende gener til å endre funksjon, ikke kan skje innen realistiske ventetider (35). Slik er mangelen på troverdighet ved neo-darwinistiske metoder som middel for å genere ny genetisk informasjon blitt påvist. Det at ventetider øker eksponentielt med økning av endog én nødvendig mutasjon, er hva vi kunne forvente ut fra Axe sin påvisning av at funksjonelle proteiner er så sjeldne. Det gjelder som helhet overfor beregninger og eksperimenter utført i perioden 2004 til 2011, at de (indirekte) bekrefter Axe sin tidligere konklusjon om sjeldenheten til funksjonelle gener og proteiner. Om neo-darwinistiske mekanismer ikke kan generere nye gener eller funksjoner for proteiner, kan de enda mye mindre bygge nye livsformer i livets tilgjengelige tidsramme på jorda.
Ved å vise det usannsynlige ved samvirke modellen for protein-evolusjon, bekreftet Axe og Gauger at gener og proteiner i seg selv representerer komplekse tilpasninger. De er avhengige av koordinert interaksjon av flere underenheter, som må oppstå som gruppe for å bevirke noen funksjonell fordel. Det innebærer igjen at biologer ikke bare kan anta at mutasjoner greit vil skape nye gener og trekk, slik det lenge har vært gjort. Både de biologer som er nevnt her, og flere andre , har nylig vist at å generere det antall multippelt-koordinerte mutasjoner som trengs endog til å produsere ett nytt gen, eller få to nærliggende gener til å endre funksjon, ikke kan skje innen realistiske ventetider (35). Slik er mangelen på troverdighet ved neo-darwinistiske metoder som middel for å genere ny genetisk informasjon blitt påvist. Det at ventetider øker eksponentielt med økning av endog én nødvendig mutasjon, er hva vi kunne forvente ut fra Axe sin påvisning av at funksjonelle proteiner er så sjeldne. Det gjelder som helhet overfor beregninger og eksperimenter utført i perioden 2004 til 2011, at de (indirekte) bekrefter Axe sin tidligere konklusjon om sjeldenheten til funksjonelle gener og proteiner. Om neo-darwinistiske mekanismer ikke kan generere nye gener eller funksjoner for proteiner, kan de enda mye mindre bygge nye livsformer i livets tilgjengelige tidsramme på jorda.
Som en ironi kan det bemerkes at det er ved å benytte verktøy som det ypperste av neo-darwinismen, populasjons-genetikken har forsynt, at en kan slutte at de neo-darwinistiske mekanismene ikke kan danne komplekse tilpasninger, inkludert nye informasjonsrike gener og proteiner, som ville være nødvendige for å bygge opp dyrene som oppsto i kambrium-perioden. For å tilpasse en metafor som Tom Frazzetta (se starten av kap. 12) ville likt: 'slangen har spist sin egen hale'.
Bilde 5. I overført betydning-har dette skjedd
Rereranser:
26. Behe og Snooke, "Simulating Evolution by Gene Duplication of Protein Features that Require Multiple Amino Acid Residues", s.2661
27. Lynch and Connery. "The Origins of Genome Complexity", s.1401-02
28. Behe og Snooke, "Simulating Evolution by Gene Duplication of Protein Features that Require Multiple Amino Acid Residues", s.2661
29. {Behe baserte seg på offentlig helse-statistikk på at det trengtes (10 opphøyd i 20 organismer) før en CCC ville inntreffe. Forutsetter en uavhengighet, vil det trenges (10 opphøyd i 40 organismer) før en dobbel CCC vil inntreffe-oversetters kommentar.}
30. M.Behe 'The Edge of Evolution', s.135
31. Dette forutsetter at alle organismene ville være under selektivt press for denne spesielle egenskapen, en totalt urealistisk forutsetning.
32. Durrett og Schmidt, "Waiting for two evolutions", 1507
33. M.Behe 'The Edge of Evolution', s.84-102
34. Gauger and Axe, 2 The Evolutionary Accessibility of New Enzyme Functions: A Case Study from the Bbiotin Pathway"
35. Gauger et al. "Reductive Evolution can Prevent Population from Taking Simple Adaptive Paths to high Fitness"; Durrett og Schmidt, "Waiting for two evolutions"
Oversettelse, samt utvalg av stoff og bilder ved Asbjørn E. Lund