Dette utviklet seg ikke ved noen få mutasjoner om gangen
Cornelius Hunter; 22. februar 2018
Oversatt herfra.
Bilde 1. GFPnevron. Kredit: Nrets på en.wikipedia [ CC BY 2.5 ], fra Wikimedia Commons.
Er det lange, gradvise, veier av funksjonelle mellomstrukturer, bare skilt av en eller kanskje noen få mutasjoner, som fører til hver enkelt art og hvert enkelt design og struktur i hele biologien? Som vi så her i går, har dette vært et grunnleggende krav og forventning innen evolusjonsteorien som er i strid med vitenskapen. Noen evolusjonister har forsøkt å komme seg rundt problemet med den nøytrale teorien, men det gjør bare saken verre.
Bilde 2. Eks. på mutasjon
 Hvis en mutasjon er sjelden, er mange mutasjoner astronomisk sjeldne. For eksempel, hvis en bestemt mutasjon har én til ett-hundre millioner (én til 10^8) sjanse for å forekomme i et nytt individ, har hundre slike spesielle mutasjoner én til (10 ^ 800) sjanse for å forekomme. Det kommer ikke til å skje. La oss ta en titt på et eksempel: nerveceller og deres potensiale for handlings-signaler.
Hvis en mutasjon er sjelden, er mange mutasjoner astronomisk sjeldne. For eksempel, hvis en bestemt mutasjon har én til ett-hundre millioner (én til 10^8) sjanse for å forekomme i et nytt individ, har hundre slike spesielle mutasjoner én til (10 ^ 800) sjanse for å forekomme. Det kommer ikke til å skje. La oss ta en titt på et eksempel: nerveceller og deres potensiale for handlings-signaler.
Nerveceller har en lang hale som bærer en elektronisk impuls. Halen kan være flere fot lang og signalet kan stimulere en muskel til handling, kontrollere en kjertel eller rapportere en følelse til hjernen.
Lik en kabel som inneholder tusenvis av forskjellige telefonledninger, blir nerveceller ofte samlet sammen for å danne en nerve. Tidlige forskere mente at kanskje den elektroniske impulsen reiste langs nervecelle-halen som strøm i en ledning. Men de skjønte snart at signalet i nerveceller er for svakt til å reise veldig langt. Nervecellen vil trenge å øke signalet underveis for å kunne forflytte seg..
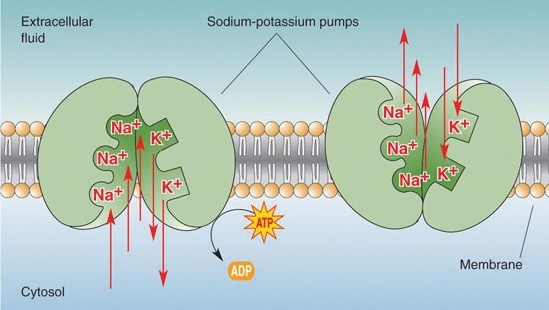
Etter mange års forskning ble det oppdaget at signalet blir forsterket av membranproteiner. For det første er det en membranprotein som samtidig pumper to kaliumioner inn i cellen og tre natriumioner ut av cellen. Dette setter opp en kjemisk gradient over membranen. Det er mer kalium inne i cellen enn ute, og det er mer natrium ute enn inne. Dessuten er det flere negativt ladede ioner inne i cellen, så det er en spenningsfall (50-100 millivolt) over membranen.
I tillegg til natrium-kaliumpumpen finnes det også natriumkanaler og kaliumkanaler. Disse membranproteinene tillater henholdsvis natrium og kalium å passere gjennom membranen. De er normalt stengt, men når den svekkede elektroniske impulsen beveger seg langs nervecellehalen, får det natriumkanaler til å åpne seg raskt. Natriumioner utenfor cellen kommer deretter til å strømme inn i cellen ned den elektrokjemiske gradienten. Som et resultat er spenningsfallet reversert, og den svekkede elektroniske impulsen, som forårsaket natriumkanalene til å åpne, økes i det den fortsetter på vei langs nervecellehalen.
Når spenningen går fra negativ til positiv i cellen, lukkes natriumkanalene seg sakte og kaliumkanalene åpnes. Derfor er natriumkanalene åpne bare kortvarig, og nå når kaliumkanalene er åpne, kommer kaliumioner som er konsentrert inne i cellen, til å strømme ut ned langs deres elektrokjemiske gradient. Som et resultat gjenopprettes den opprinnelige spenningsforskjellen.
Denne prosessen gjentar seg selv da den elektroniske impulsen beveger seg langs nervecellen, til impulsen endelig når slutten av nervecellen. Selv om vi har utelatt mange detaljer, bør det være åpenbart at prosessen avhenger av de intrikate virkemåtene til de tre membranproteinene. Natriumkaliumpumpen bidrar til å opprette den elektrokjemiske gradienten, den elektroniske impulsen er sterk nok til å aktivere natriumkanalen, og deretter åpnes og lukker natrium- og kaliumkanaler med presis timing.
Bilde 3. Eks. på ione-kanal
 Hvordan, kan det for eksempel, ha seg at kanalene er designet for å være ion-selektive? Natrium er ca 40 prosent mindre enn kalium, slik at natriumkanalen kan utelukke kalium hvis den er akkurat stor nok til natrium. Tilfeldige mutasjoner må ha slått på en aminosyresekvens som vil folde seg akkurat korrekt for å gi riktig kanalstørrelse.
Hvordan, kan det for eksempel, ha seg at kanalene er designet for å være ion-selektive? Natrium er ca 40 prosent mindre enn kalium, slik at natriumkanalen kan utelukke kalium hvis den er akkurat stor nok til natrium. Tilfeldige mutasjoner må ha slått på en aminosyresekvens som vil folde seg akkurat korrekt for å gi riktig kanalstørrelse.
Kaliumkanalen er derimot stor nok til både kalium og natrium, men den er svært effektivt. Det eliminerer natrium på et vis nesten perfekt (kalium til natrium-forholdet er ca. 10.000), men tillater at kalium passerer nesten som om det ikke var noe i veien.
Nerveceller virker kontinuerlig i kroppen din. De kontrollerer øynene dine mens du leser disse ordene, og de sender tilbake bildene du ser på denne siden til hjernen din. De styrer, sammen med kjemiske signaler, en rekke prosesser i kroppene våre, og det er ingen vitenskapelig grunn til å tro at de gradvis utviklet seg, med én mutasjon om gangen.
Faktisk er denne ideen i strid med alt vi vet fra vitenskapen. Og likevel tror evolusjonistene dette. La meg gjenta at evolusjonister tror at nerveceller og deres handlingspotensiale-design utviklet seg med én mutasjon om gangen. Faktisk tror evolusjonister at dette er et bevist faktum, utover all rimelig tvil -sml. tyngdekraften.
Det ville være vanskelig å forestille seg et mer absurd krav. Så la oss ta en titt på detaljene i denne tankegangen. Her er en nylig artikkel fra Royal Society, som representerer status i evolusjonell tenkning om dette emnet. Artikkelen hevder å gi en detaljert forklaring på hvor tidlig evolusjon produserte handlingspotensiell teknologi.
Høres lovende, men når evolusjonister snakker om “detaljer”, har de noe litt annet i tankene:
Bilde 4. Standard nålevende amøbe
 Her er flere passasjer fra artikkelen {innledes med -- som så kommenteres punkt for punkt -oversetters tilføyelse}som viser at det ikke bare er mangel på detaljer, men at studien er grundig uvitenskapelig:
Her er flere passasjer fra artikkelen {innledes med -- som så kommenteres punkt for punkt -oversetters tilføyelse}som viser at det ikke bare er mangel på detaljer, men at studien er grundig uvitenskapelig:
--Vi foreslår at neste trinn i utviklingen av eukaryot DCS [membran depolarisering (ved ukontrollert kalsium tilstrømning), sammentrekning og sekresjon] kopling har vært rekruttering av strekkfølsomme kalsiumkanaler, som tillater kontrollert tilstrømning av kalsium ved mekanisk stress før selve skaden oppstår, og dermed forutse virkningene av membranbrudd.
Rekruttering av kalsiumkanaler? Og hvem nøyaktig rekrutterte dem? Her stoler forfatterne på vag terminologi for å dekke over en rekke problematiske detaljer om hvor tilfeldige mutasjoner på en eller annen måte utførte denne rekruttering.
--For å forhindre selve bruddet kunne den første rollen som mekano-sensoriske Ca ++-kanaler ha vært å forhåndsaktivere komponenter i reparasjonsveien i strekte membraner.
"å forhindre"? La oss male ut logikken litt tydeligere. Forfatterne hypotetiserer at disse kalsiumkanalene utviklet muligheten til å preaktivere reparasjonsveien “for å forhindre” faktisk brudd. Ved å male ut logikken litt tydeligere, kan vi lettere se den vanlige teleologien på ferde. Evolusjonslitteraturen er full av teleologi, og med god grunn. Evolusjonister kan ikke formulere og uttrykke sine ideer uten det. Den evigvarende infinitive form er et megetsigende tegn. Aristotelianismen er død – lenge leve aristotelianismen.
Bilde 5. Prinsippskisse av nervecelle
 --Som et annet forventet tiltak , kan actomyosin ha vært forhånds-posisjonert under plasmamembranen (derav det kortikale actomyosin-nettverket oppdaget i hver eukaryotisk celle) og kan også ha utviklet direkte følsomhet for strekking.. Når dets kortikale posisjon og mekano-sensibilitet ble ervervet, kunne actomyosin-nettverket automatisk oppfylle en tilleggsfunksjon : Vedlikehold av celleformer - da lokalisert vil celledeformasjon strekke det kortikale actomyosin-nettverket og utløse en umiddelbar kompenserende sammentrekning. Denne egenskapen ville ha oppstått som en bivirkning .. av tilstedeværelsen av kortikal actomyosin for membranreparasjon, og viste seg raskt å være fordelaktig.
--Som et annet forventet tiltak , kan actomyosin ha vært forhånds-posisjonert under plasmamembranen (derav det kortikale actomyosin-nettverket oppdaget i hver eukaryotisk celle) og kan også ha utviklet direkte følsomhet for strekking.. Når dets kortikale posisjon og mekano-sensibilitet ble ervervet, kunne actomyosin-nettverket automatisk oppfylle en tilleggsfunksjon : Vedlikehold av celleformer - da lokalisert vil celledeformasjon strekke det kortikale actomyosin-nettverket og utløse en umiddelbar kompenserende sammentrekning. Denne egenskapen ville ha oppstått som en bivirkning .. av tilstedeværelsen av kortikal actomyosin for membranreparasjon, og viste seg raskt å være fordelaktig.
Et "foregående trinn"? "Pre-posisjonering"? Actomyosin "utviklet" følsomhet for strekk? Stillingen og mekanosensibiliteten "ble ervervet"? Nettverket kan "oppfylle en ekstra funksjon"? Beklager, men molekylære maskiner (for eksempel actomyosin) "utvikler ikke" noe. Det er mer teleologi pakket inn i disse få setningene enn i noen middelalderlig pamflett. Og for et godt mål tilføyer forfatterne også det forbløffende lykketreff at denne tilleggsfunksjonen "ville ha oppstått som en bivirkning." Det var heldig.
Bilde 6. ‘Cellens lim’ -laminin
 --Når den dekket celle-barken, ervervet actomyosin-nettverket evnen til å deformere cellen ved lokalisert sammentrekning.
--Når den dekket celle-barken, ervervet actomyosin-nettverket evnen til å deformere cellen ved lokalisert sammentrekning.
Actomyosin-nettverket: "ervervet evnen" til å deformere cellen ved lokalisert sammentrekning? Smart trekk på vegne av nettverket. Men kan vi spørre hvordan skjedde det?
--Basert på den genomiske studien av den protistiske (lenke) Naegleria som har en bifasisk livssyklus (alternerende mellom en amøbisk og en flagellert fase), er amøbisk-fremdrift blitt foreslått å være stamfar for eukaryoter. Det kan ha utviklet seg i begrensede mellom-miljøer, da det er spesielt instrumentalt for celler som trenger å bevege seg gjennom små, uregelmessig formede rom ved utforskende deformasjon.
Amøbisk forflytning utviklet seg "etter som det er spesielt instrumentelt." Ingen infinitiv form, men dette er ikke mindre teleologisk. Ting utvikler seg ikke fordi de er "instrumentale". Forfatterne unnlater å informere sine lesere om at dette ville kreve et enormt antall tilfeldige mutasjoner.
--Man kan hypotetisere at hvis strekningskjenslige kalsiumkanaler og kortikal actomyosin var en del av den eukaryote molekylære verktøykassen til stamfedre (som sammenlignende genomikk –her indikerer), ville membran deformasjon i et begrenset miljø trolig utløse kalsium tilstrømning ved å åpne strekkfølsomme kanaler, noe som i sin tur ville fremkalle bred actomyosin sammentrekning over den deformerte delen av cellebarken, global deformasjon og cellebevegelse bort fra trykkilden.
Konseptet med en "molekylær verktøykasse" er standard i evolusjonær tanke, og et annet eksempel på teleologisk tenkning.
--Man kan således foreslå at en enkel forfedreform for amøbeaktig bevegelse utviklet seg som en naturlig konsekvens av scenariet som er skissert ovenfor for opprinnelsen til kortikal actomyosin og kalsium-kontraksjonskoblingen; når det først var etablert, kunne det ha blitt videreutviklet.
Amøbe-bevegelsen utviklet seg som en naturlig konsekvens, og "en gang etablert" ble det "videreutviklet"? Dette er ikke noe annet enn teleologisk historiefortelling uten støttende bevis.
Bilde 7. Pilin-flagell

--Det er derfor fristende å spekulere på at når kalsium-signalering hadde fått kontroll over primitive former for amøbebasert bevegelse, begynte det samme signalsystemet å modifisere cilias kraftslag, muligens for å svitsje mellom bevegelige tilstander.
Kalsium signalering “fikk kontroll” og begynte deretter “å modifisere” cilias kraftslag “for”å svitsje “mellom bevegelsesaktige tilstander”? “For å svitsje” er enda et uttrykk velduftende av teleologi, og “vant kontroll” er et aktivt trekk ved kalsium-signalanlegget. Ren, uforfalsket, teleologi.
--Kanskje induserte kalsium i forfedre eukaryoter en relativt enkel bryter (som ciliær arrest, slik den fremdeles ses i mange dyreceller og i Chlamydomonas som respons på høye Ca++ konsentrasjoner), som deretter ble gradvis endret til mer subtile moduleringer av slagmodus med en rask omsetning av molekylære aktører formidlet ved differensialt tillegg, komplementering og tap.
"Kalsium induserte en relativt enkel bryter"? Beklager, ioner forårsaker ikke brytere, enkle eller ikke. Og bryteren “ble deretter gradvis endret til mer subtile moduleringer”? Legg merke til hvordan den passive stemmen fjerner de ekle detaljene. Bryteren “ble modifisert” utelater praktisk det faktum at slik modifikasjon måtte oppstå via tilfeldige mutasjoner, én mutasjon av gangen.
 --Alternativt kunne kontroll av cilia ved kalsium ha utviklet seg konvergent –lenke, men en slik konvergens ville da vært bemerkelsesverdig allestedsnærværende, da det ikke synes å være noe eukaryotisk flagellum som ikke er kontrollert av kalsium på en eller annen måte.
--Alternativt kunne kontroll av cilia ved kalsium ha utviklet seg konvergent –lenke, men en slik konvergens ville da vært bemerkelsesverdig allestedsnærværende, da det ikke synes å være noe eukaryotisk flagellum som ikke er kontrollert av kalsium på en eller annen måte.
"Kunne ha utviklet seg konvergent"? Og hvordan nøyaktig ville det skje? I det minste innrømmer forfatterne absurditeten i det alternativet.
Dessverre mangler de slik følsomhet for resten av artikkelen. Som vi så over, er artikkelen basert på en sekvens av teleologisk tenkning. Det faller inn i den evolusjonære sjangeren der evolusjon er tatt, a priori, som gitt. Dette går i antagelse underskriver enorme strekk av teleologisk tanke, og tegneserie-nivå historie-fortelling. Ikke bare er det mangel på empirisk støtte, men sjangeren er helt uvitenskapelig, som åpenbart ved selv en mild kritisk lesning.
Og unødvendig å si, artikkelen gjør absolutt ingenting for å lindre problemet vi begynte med. De mange sprangene av logikk og resonnement i artikkelen avslører alle slags monumentale forandringer evolusjon krever ‘for å’ konstruere nerveceller og handlingspotensiell teknologi. Vi ser ikke på en fortelling om små, gradvise endringer som hver bidrar til den generelle egnetheten. Mange, mange samtidige mutasjoner kommer til å trengs. Selv et konservativt minimums antall på 100 samtidige mutasjoner fører til det uholdbare resultatet av én til (10^800) sjanse for å inntreffe.
Det kommer ikke til å skje.
Oversettelse ved Asbjørn E. Lund