 Liv eksponentielt: Livet viser intelligent design på mange nivåer
Liv eksponentielt: Livet viser intelligent design på mange nivåer
Jonathan Wells; 1. juni 2018
Oversatt herfra.
Intelligent design
Bilde 1. Kredit: Froske egg, by Geoff Gallice from Gainesville, FL, USA [CC BY 2.0], via Wikimedia Commons.
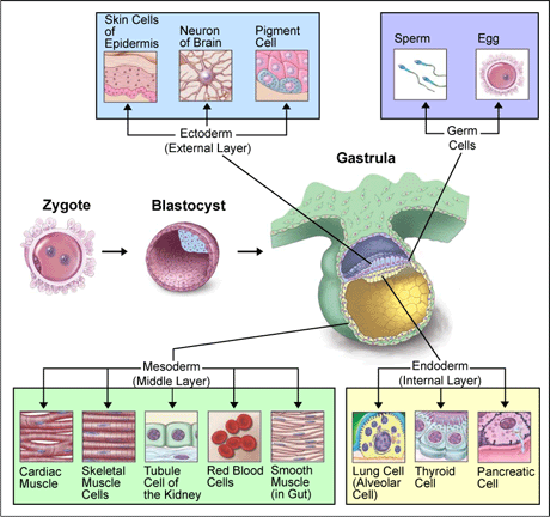
En menneskelig celle inneholder to sett med DNA, som hver består av om lag tre milliarder underenheter kalt "nukleotider." Det er fire forskjellige nukleotider, og de kan ordnes på mange forskjellige måter, så DNA er ganske komplekst. Det meste av vårt DNA må imidlertid ordnes på en svært spesifikk måte for å gi informasjonen en celle bruker til å lage RNA og proteiner. Matematiker William Dembski har kalt denne "kompleks spesifisert informasjon." (1)
Kompleksitet (som vi ser i en haug av høstblader) kan oppstå spontant fra ikke-styrte naturlige prosesser, men kompleks spesifisert informasjon kan ikke. Den eneste kjente kilden til kompleks spesifisert informasjon er et intelligent sinn som kan forestille seg et mål og ordne ting for å realisere det målet - i dette tilfellet en levende celle. "Fordi sinn eller intelligent design er en nødvendig årsak til et informasjonsrikt system, skrev forsknings-filosofen Stephen Meyer i 1998," de spesifikt arrangerte nukleotidsekvensene - den kodede informasjonen - i DNA indikerer tidligere handling til et intelligent sinn, til og med om ikke en slikt mental agent kan observeres direkte. " (2)
Behovet for romlig informasjon
Så DNA bærer biologisk informasjon, og informasjon peker på design. Men DNA er ikke den eneste bærer av informasjon i en levende celle. Når DNA transkriberes til RNA, må de fleste av disse RNA'ene transporteres til bestemte steder i cellen før de kan fungere skikkelig. Noen RNA er merket med sekvenser kalt "zip-koder" som angir "adressene" som de skal transporteres til. Som postnummeret på et brev du legger inn en postboks, er imidlertid et RNA-postnummer meningsløs med mindre den tilsvarer en eksisterende adresse. I likhet med de geografiske adressene i et postsystem, må de mobile destinasjonene til RNA og proteiner spesifiseres uavhengig av deres postnummer, før de blir "sendt".
Mange av disse destinasjonene er spesifisert av molekyler innebygd i cellemembraner. Forskere opprinnelig trodde at proteiner kunne diffundere fritt i en membran, som båter som flyter på sjøen. Det er imidlertid kjent at mange membranproteiner er 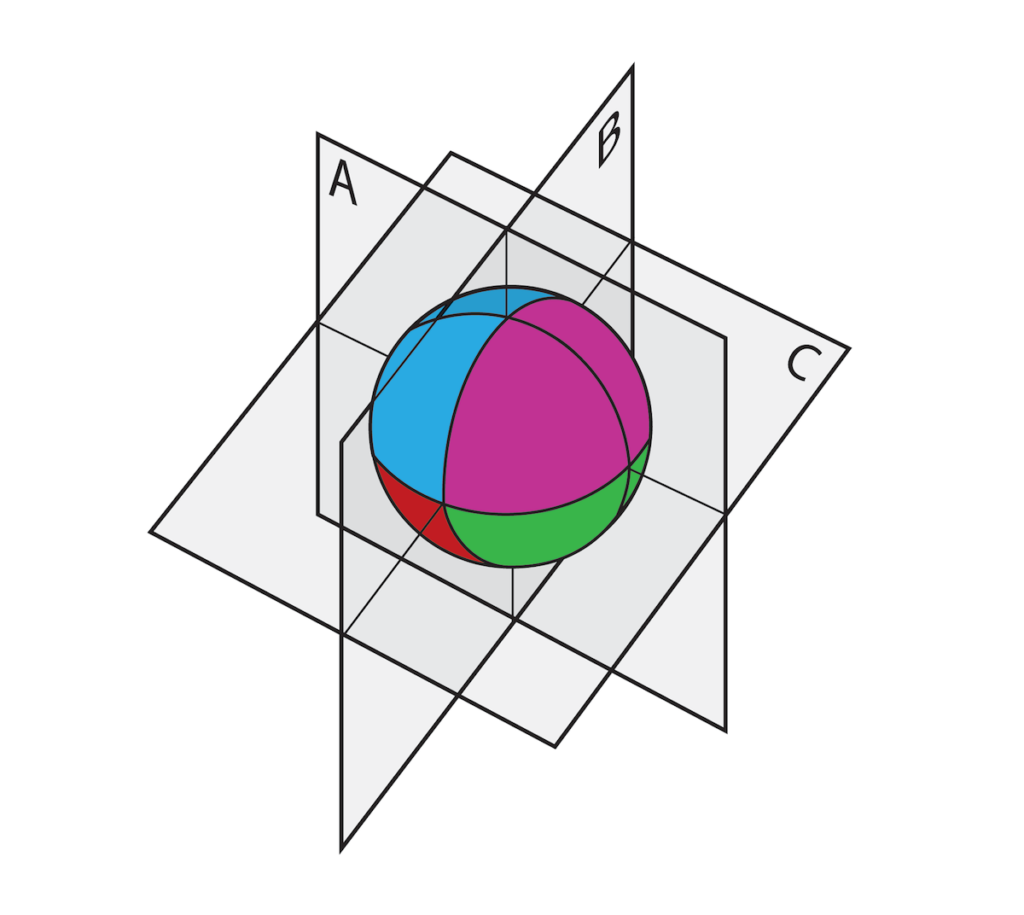 arrangert i ikke-tilfeldige mønstre som kan være ganske stabile. Slike mønstre forsyner cellen med romlig informasjon som går utover informasjonen i DNA.
arrangert i ikke-tilfeldige mønstre som kan være ganske stabile. Slike mønstre forsyner cellen med romlig informasjon som går utover informasjonen i DNA.
Etter befruktning er et froske-embryo usynlig delt inn i regioner som er preget av (blant annet) romlig lokaliserte RNAer like innenfor cellemembranen i en sone som kalles "cortex". I den medfølgende tegningen er fire områder indikert med forskjellige farger. Etter at kjernen dupliserer, deles det befruktede egget i to datterceller. Hvis divisjonsplanet tilsvarer A, arver hver dattercelle ikke bare en kjerne, men også deler av alle fire regionene av kortikal informasjon. Hvis de to cellene separeres, kan hver utvikle seg til en komplett frosk. Men hvis divisjonsplanet tilsvarer B eller C, arver ikke dattercellene et fullt sett av kortikal informasjon, og deres utvikling er blokkert, selv om de hver inneholder alt nødvendig DNA.
Illustrasjon: Regioner av romlig informasjon i et befruktet froskeegg.
Andre transportører av romlig informasjon
Regionale forskjeller i celler og embryoer kan spesifiseres på andre måter, i tillegg til lokalisering av RNA i cortex. To av disse måtene har blitt studert i stor detalj: "sukkerkoden" og den "bioelektriske koden".
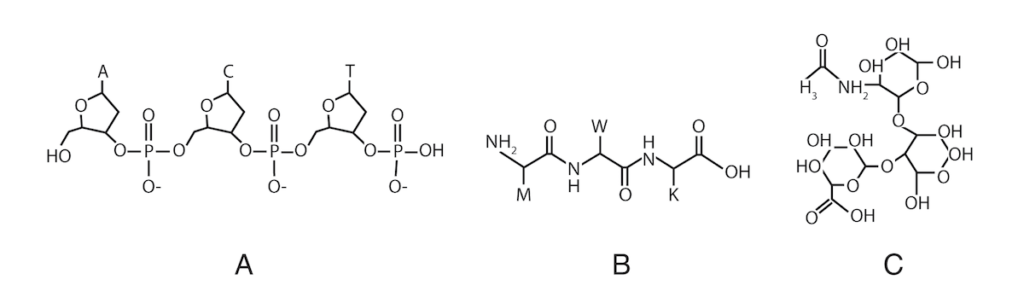
De fleste proteiner i levende celler - inkludert de i membraner - er kjemisk bundet til karbohydrater kalt "glykaner" (fra det greske ordet for "søtt"). Nukleotidene i DNA er koblet sammen ende-til-ende i et lineært molekyl, så DNA-sekvensinformasjon er endimensjonal. I levende celler er underenhetene i proteiner (med noen få unntak) også forbundet i en lineær kjede. Men glykaner kan kobles sammen på komplekse tredimensjonale måter, slik at deres informasjonskapasitet overstiger DNA og proteiner med mange størrelsesordener. (3)
 Informasjonen som bæres av glykaner har blitt kalt "sukkerkoden". (4) Sukkerkoden er "tolket" av proteiner som kalles lektiner, som "gjenkjenner" spesifikke tredimensjonale strukturer av glykanmolekyler. Glykaner og lektiner spiller en viktig rolle i kommunikasjon mellom celler og bidrar til å veilede cellebevegelser i et utviklende embryo. Eksperimenter har vist at membranmønstre av glykaner endres i løpet av embryoutvikling. (5)
Informasjonen som bæres av glykaner har blitt kalt "sukkerkoden". (4) Sukkerkoden er "tolket" av proteiner som kalles lektiner, som "gjenkjenner" spesifikke tredimensjonale strukturer av glykanmolekyler. Glykaner og lektiner spiller en viktig rolle i kommunikasjon mellom celler og bidrar til å veilede cellebevegelser i et utviklende embryo. Eksperimenter har vist at membranmønstre av glykaner endres i løpet av embryoutvikling. (5)
Illustrasjon: Tre underenheter knyttet sammen i DNA (A), et protein (B) og et glykan (C).
I tillegg til sukkerkoden, genererer sannsynligvis alle levende celler (ikke bare nerve- og muskelceller) elektriske felt over deres membraner. De gjør dette ved å pumpe ladde ioner gjennom kanaler i membranene deres, og skape et "bioelektrisk felt". Mønsteret av membrankanaler bestemmer form av det bioelektriske feltet, og formen på feltet endres under embryoutvikling.
Illustrasjon: Noen av de bioelektriske feltene (vist med piler) i et utviklende froskembryo.
 Bioelektriske felt er korrelert med viktige utviklingshendelser. I froskembryoer begynner for eksempel store ione-strømmer å strømme ut av de stedene hvor baklemmen vil utvikle seg lenge før lemmerne faktisk opptrer.
Bioelektriske felt er korrelert med viktige utviklingshendelser. I froskembryoer begynner for eksempel store ione-strømmer å strømme ut av de stedene hvor baklemmen vil utvikle seg lenge før lemmerne faktisk opptrer.
Mange eksperimenter utført siden 1980-tallet har bekreftet at forstyrrende bioelektriske felt forårsaker forstyrrelser i utviklingen. For eksempel genererer froskembryoer normalt et elektrisk felt i hode til hale retning. Hvis et kunstig felt av samme størrelsesorden blir brukt i motsatt retning, eller hvis ionekanaler som genererer feltet, er blokkert, er resultatet unormal utvikling av hode og øyne. Stedene hvor øynene normalt dannes er mer høyt ladet enn det omgivende vevet; Hvis ladningen er nøytralisert, er øynene som utvikles små eller deformerte. Noen ganger utvikler øyne seg andre steder på rumpetrollets kropp, inkludert halen. (5)
Hvordan påvirker elektriske felt utvikling? På 1980-tallet eksponerte biologer embryonale celler til kunstige elektriske felt med samme styrke som cellene genererer naturlig. Noen typer celler migrerte mot den positive polen, mens andre typer migrerte til den negative polen, noe som tyder på at enveis bioelektriske felter påvirker embryoutvikling ved å styre cellebevegelser.
I 1995 konkluderte biologene Riyi Shi og Richards Borgens at bioelektriske felt "kan forsyne et tredimensjonalt koordinatsystem" som bidrar til å spesifisere form i embryoer. (6) I 2013 skrev biologer AiSun Tseng og Michael Levin at slike felt kan gi "form-maler", og at en full forståelse av embryoutvikling sannsynligvis vil kreve å knekke den "bioelektrisk koden" (7) .
 Membran-koden
Membran-koden
Så, lokaliserte RNA i cortex, glykanmønstre på membranen og bioelektriske felt som genereres av ionekanaler i membranen, har alle romlig informasjon. Selv om enkelte molekyler kan spesifiseres av DNA, blir ikke deres tredimensjonale mønstre det. Samlet sett utgjør disse mønstrene en "membrankode" som er uavhengig av DNA-sekvenser.
I 1983 foreslo biolog Robert Poyton at biologiske membraner bærer "romlig minne", hvor enhetene er romlig lokaliserte proteiner. Poyton skrev: "å realisere at genetisk minne er endimensjonalt, langs et DNA-molekyl, mens romlig hukommelse sannsynligvis er todimensjonal, langs membranflater og tredimensjonal innenfor det cellulære interiøret, er det sannsynlig at romlig minne er mer komplisert og mangfoldig enn genetisk minne." (8)
I 2004 skrev biolog Thomas Cavalier-Smith at ideen om at DNA inneholder all den informasjonen som trengs for å lage en organisme "er rett og slett falsk." Ifølge Cavalier-Smith, gir membraner "kjemisk spesifikke todimensjonale overflater med gjensidig konserverte topologiske relasjoner i de tre romlige dimensjonene som spiller en nøkkelrolle i mekanismene som konverterer lineær informasjon av DNA til tredimensjonale former for enkeltceller og multicellulære organismer. Dyreutvikling skaper en kompleks tredimensjonal multicellulær organisme, ikke ved å starte fra den lineære informasjonen i DNA ... men starter alltid fra en allerede svært komplisert tredimensjonal encellet organisme, det befruktede egget." (9)
Så membrankoden bærer viktig biologisk informasjon som er uavhengig av DNA-sekvensinformasjon. Likevel hører vi ofte at embryo-utvikling styres av et program i DNA. Hvorfor?
Bilde. Embryo-utvikling ikke fullstendig DNA-styrt
Utover DNA
James Watson og Francis Cricks Nobelprisvinnende oppdagelse av DNAs molekylære struktur i 1953 syntes å gi en molekylær basis, ikke bare for arvelighet, men også for embry-outvikling. Celler replikerer deres DNA før de deles og sender (vanligvis) et komplett sett av deres DNA-sekvenser til hver av sine etterkommere. Celler bruker da DNA-sekvenser som maler for transkripsjon av RNA, hvorav noen deretter blir oversatt til proteiner.
På midten av 1900-tallet ble biologi dominert av neo-darwinisme, et tanke-system som kombinerte evolusjon og genetikk og tilskrev nye variasjoner til genetiske mutasjoner. En underliggende forutsetning for neo-darwinismen er at utvikling bare skyldes ikke-styrte materielle prosesser. Etter 1953 førte denne materialistiske antagelsen til at "DNA lager RNA som lager protein som lager oss", som har blitt kalt Sentral-dogmet i molekylær biologi.
I 1970 sa molekylær-biologen (og materialistiske) Jacques Monod at med det sentrale dogmet "og forståelsen av det tilfeldige fysiske grunnlaget for mutasjoner som molekylærbiologi også har gitt, er darwinismens mekanisme endelig sikkert grunnlagt. Og mennesket må forstå at han bare er en ulykke." (9)
Men eksistensen av membrankoden viser at det sentrale dogmet er feilaktig. Og den materialistiske ideen om at evolusjonen er ikke-styrt, ikke kan gjøre rede for den komplekse spesifiserte informasjonen i DNA, mye mindre for den omfattende komplekse spesifiserte informasjonen i membrankoden. Akkurat som informasjonen i DNA peker på design, gjør også informasjonen utover DNA det.
 Merknader :
Merknader :
1. William A. Dembski, "Intelligent design as a theory of information", February 20, 1997.
2. Stephen C. Meyer, "DNA by design", Rhetoric and Public Affairs 1 (1998): 519-556.
3. Roger A. Laine, "A calculation of all possible oligosaccharide isomers both branched and linear yields 1.05 x 1012 structures for a reducing hexasaccharide", Glycobiology 4 (1994): 759-767.
4. Hans-Joachim Gabius, "Biological information transfer beyond the genetic code: The sugar code", Naturwissenschaften 87 (2000): 108-121.
5. Jonathan Wells, "Membrane patterns carry ontogenetic information that is specified independently of DNA", Bio-Complexity 2014 (2): 1-28.
6. Riyi Shi and Richard B. Borgens, "Three-dimensional gradients of voltage during development of the nervous system as invisible coordinates for the establishment of embryonic pattern", Developmental Dynamics 202 (1995): 101-114.
7. AiSun Tseng and Michael Levin, "Cracking the bioelectric code: Probing endogenous ionic controls of pattern formation", Communicative and Integrative Biology 6 (2013): e22595
8. Robert O. Poyton, "Memory and membranes: The expression of genetic and spatial memory during the assembly of organelle macrocompartments", Modern Cell Biology 2 (1983): 15-72.
9. Thomas Cavalier-Smith, "The membranome and membrane heredity in development and evolution", pp. 335-351 in Robert P. Hirt and David S. Horner (editors), Organelles, Genomes and Eukaryote Phylogeny (Boca Raton, FL: CRC Press, 2004), 348.
10. Jacques Monod, quoted in Horace Freeland Judson, The Eighth Day of Creation: The Makers of the Revolution in Biology (New York: Simon & Schuster, 1979), 217.
Editor's note: Dr. Wells' latest book is "Zombie Science: More Icons of Evolution". This article first appeared in Salvo 38. It is published here with the permission of Jonathan Wells.
Oversettelse og bilder ved Asbjørn E. Lund