Stephen Meyers omfattende behandling av Evo-devo
Av Casey Luskin, 24. juli 2024. Oversatt herfra
 I går -lenke bemerket jeg at Gerd Müller -lenke, emeritus -professor ved University of Wien, i fjor hadde svart på Stephen Meyerss poeng fra Joe Rogan -podcasten om utvikling og evolusjon. Müller er kritiker av neo-darwinsk teori og dens evne til å forklare opprinnelsen til biologisk nyhet, og foretrekker i stedet Evo-devo-baserte modeller for å forklare hvordan nye kroppsplaner oppstår. Selv om dette ikke kom mye opp i Rogan-podcasten, skrev Meyer i Darwins Doubt faktisk ganske mye om Evo-devo-baserte modeller av organisme evolusjon.
I går -lenke bemerket jeg at Gerd Müller -lenke, emeritus -professor ved University of Wien, i fjor hadde svart på Stephen Meyerss poeng fra Joe Rogan -podcasten om utvikling og evolusjon. Müller er kritiker av neo-darwinsk teori og dens evne til å forklare opprinnelsen til biologisk nyhet, og foretrekker i stedet Evo-devo-baserte modeller for å forklare hvordan nye kroppsplaner oppstår. Selv om dette ikke kom mye opp i Rogan-podcasten, skrev Meyer i Darwins Doubt faktisk ganske mye om Evo-devo-baserte modeller av organisme evolusjon.
Blde 1. Kråkebolle
Sett deg for en lang lesning
Forbered deg på en lang lesning fordi i boka bruker Meyer mye plass til emnet. Dette er hva han skriver:
Evolusjonsteoretikere og utviklingsbiologer som Rudolf Raff, Sean B. Carroll og Wallace Arthur har utviklet en subdisiplin av biologi, kjent som evolusjonær utviklingsbiologi, eller kortformen "evo-devo". De evolusjonære utviklings-biologene har siden formulert alternative modeller som utfordrer et sentralt aspekt av den neo-darwiniske triaden. Mens neo-darwinisme ser for seg ny form som oppstår som et resultat av langsomme, trinnvise ansamlinger av mindre mutasjoner, hevder evolusjons-utviklingsbiologer at mutasjoner som påvirker gener som er involvert i dyreutvikling, kan forårsake storskala morfologisk endring og til og med helt nye kroppsplaner .. En tilnærming faller under rubrikken til "Evo-Devo" og forestillinger om mutasjoner som produserer modifikasjoner i større trinn.
Evo- Devo og dens forslag
Den neo-darwinske syntesen har lenge lagt vekt på at storskala makroevolusjonær endring skjer som det uunngåelige biproduktet av akkumulering av småskala "mikroevolusjonære" endringer i populasjoner. Konsensus til støtte for denne ideen begynte å frynses opp i evolusjonsbiologi på begynnelsen av 1970-tallet, da unge paleontologer som Gould, Niles Eldredge og Steven Stanley innså at fossilprotokollen ikke viste et mønster av gradvis "mikro-til-makro" endring. I 1980, på et nå kjent symposium om makroevolusjon på Field Museum i Chicago, kom opprøret fullt til syne, og avslørte det utviklingsbiolog Scott Gilbert kalte "en underjordisk strøm i evolusjonsteori" blant teoretikere som hadde konkludert med at "makroevolusjon ikke kunne være avledet fra mikroevolusjon. "
Bilde 2. Tvilende til mikro-makro konsensus (Lynn Margulis)
 På konferansen fant paleontologer som tvilte på "mikro-til-makro" -konsensus allierte blant yngre utviklingsbiologer. De var misfornøyde med neo-darwinisme delvis fordi de visste at populasjonsgenetikk, dets matematiske uttrykk, bare søkte å kvantifisere endringer i genfrekvens i stedet for å forklare opprinnelsen til gener eller nye kroppsplaner. Dermed trodde mange utviklingsbiologer at neo-darwinisme ikke ga en overbevisende teori om makroevolusjon.
På konferansen fant paleontologer som tvilte på "mikro-til-makro" -konsensus allierte blant yngre utviklingsbiologer. De var misfornøyde med neo-darwinisme delvis fordi de visste at populasjonsgenetikk, dets matematiske uttrykk, bare søkte å kvantifisere endringer i genfrekvens i stedet for å forklare opprinnelsen til gener eller nye kroppsplaner. Dermed trodde mange utviklingsbiologer at neo-darwinisme ikke ga en overbevisende teori om makroevolusjon.
For å formulere en mer robust teori, oppfordret mange utviklingsbiologer, som Rudolf Raff, en utviklingsbiolog ved University of Indiana og en av grunnleggerne av "Evo-Devo", evolusjonsteoretikere til å innlemme innsikt fra disiplinen. For eksempel vet utviklingsbiologer at mutasjoner uttrykt tidlig i utviklingen av dyr er nødvendige for å endre kroppsplan-morfogenese. Dermed hevder de at disse mutasjonene må ha spilt en betydelig rolle i å generere helt nye dyreformer under livets historie. De hevder at denne forståelsen av utviklingsprosesser er avgjørende for å forstå dyreutviklingen. Noen talsmenn for Evo-DeVo som Sean B. Carroll og Jeffrey Schwartz har spesifikt pekt på homeotiske (eller Hox) gener- master regulatoriske gener som påvirker plasseringen, timingen og uttrykk for andre gener- som enheter som er i stand til å produsere storskala endring i dyreform. Disse Evo-Devo talsmenn har brutt med klassisk neo-darwinisme først og fremst i deres forståelse av størrelsen eller økningen av mutasjonsendring.
Stor, men ikke levedyktig, levedyktig, men ikke stor
Til tross for entusiasmen rundt feltet, mislykkes Evo-devo, og av en åpenbar grunn: Dets viktigste forslag, at tidligvirkende utviklingsmutasjoner kan forårsake stabilt arvelige, store endringer i dyre kroppsplaner, motsier resultatene fra hundre år med Mutagenese eksperimenter. Som vi så i kapittel 13, har eksperimentene til forskere som Nüsslein-Volhard og Wieschaus definitivt vist at tidligvirkende kroppsplanmutasjoner alltid genererer embryonalt døde dyr som ikke er i stand til videre evolusjon. Resultatene fra disse eksperimentene har generert dilemmaet for evolusjonsbiologer som genetikeren John McDonald passende beskrev som det "store darwinistiske paradokset." Husk at McDonald bemerket at tidligvirkende regulatoriske mutasjoner ikke gir levedyktige endringer i form som vil vedvare i populasjoner, slik evolusjonen absolutt krever. I stedet elimineres disse mutasjonene umiddelbart ved naturlig seleksjon på grunn av deres alltid ødeleggende konsekvenser. På den annen side kan senere virkende mutasjoner generere levedyktige endringer i funksjonene til dyr, men disse endringene påvirker ikke globale dyrearkitekturer. Dette genererer et dilemma: Store endringer er ikke levedyktige; levedyktige endringer er ikke store. I ingen av tilfellene gir den typen mutasjon som faktisk forekommer levedyktige store endringer av den typen som er nødvendige for å bygge nye kroppsplaner.
I 2007 var jeg medforfatter til en lærebok med flere kolleger med tittelen Explore Evolution. I den forklarte vi dette "enten/eller" ("major-ikke-levedyktig, levedyktig-ikke-major") dilemma og antydet at det utgjorde en utfordring for teorier som er avhengige av mutasjons- og seleksjonsmekanismen for å forklare opprinnelsen til store morfologiske endringer. National Center for Science Education (NCSE) - en innflytelsesrik aktivistgruppe som motsetter seg at studentene kan lære om vitenskapelig kritikk av evolusjonsteori - utfordret kritikken vår. De påsto at læreboka vår "ikke klarer å erkjenne den omfattende forskningen på mutasjoner i DNA -sekvenser som ikke koder for proteiner, men som har viktige morfologiske effekter." Med andre ord hevdet de at noen levedyktige mutasjoner gir store storskala endringer.
 NCSE siterte artikler fra "Evo-Devo" -litteraturen og hevdet at en type mutasjon i reguleringsregionene i genomet, "cis-regulerende" regioner, har vist seg å produsere store endringer i bevingede insekter. I følge NCSE blir mutasjoner i disse Cis-Regulerende Elementene (eller CRE) ansett av mange evolusjonsbiologer til å ha det største potensialet for å generere evolusjonsendring. " Dessuten insisterte de på at "mutasjoner i CRE spiller en viktig rolle i morfologisk evolusjon." NCSE siterte en artikkel i Proceedings of the National Academy of Sciences av tre utviklingsbiologer, Benjamin Prud’homme, Nicolas Gompel og Sean B. Carroll. Oppgaven viste ikke hva NCSE hevdet. Den hevdet at endringer i regulatorisk DNA produserer "både relativt beskjedne morfologiske forskjeller mellom nært beslektede arter og dypere anatomiske avvik mellom grupper på høyere taksonomiske nivåer." Men studien viste bare hvordan endringer i de cis-regulerende elementene i fruktflue-DNA kan ha påvirket fargen på vingestedene i flere forskjellige typer flygende insekter. Den rapporterte ingen vesentlig endring i form eller kroppsplan for disse insektene. I stedet fremhevet studien et tydelig tilfelle av en levedyktig mutasjon som bare genererer en mindre eller overfladisk endring.
NCSE siterte artikler fra "Evo-Devo" -litteraturen og hevdet at en type mutasjon i reguleringsregionene i genomet, "cis-regulerende" regioner, har vist seg å produsere store endringer i bevingede insekter. I følge NCSE blir mutasjoner i disse Cis-Regulerende Elementene (eller CRE) ansett av mange evolusjonsbiologer til å ha det største potensialet for å generere evolusjonsendring. " Dessuten insisterte de på at "mutasjoner i CRE spiller en viktig rolle i morfologisk evolusjon." NCSE siterte en artikkel i Proceedings of the National Academy of Sciences av tre utviklingsbiologer, Benjamin Prud’homme, Nicolas Gompel og Sean B. Carroll. Oppgaven viste ikke hva NCSE hevdet. Den hevdet at endringer i regulatorisk DNA produserer "både relativt beskjedne morfologiske forskjeller mellom nært beslektede arter og dypere anatomiske avvik mellom grupper på høyere taksonomiske nivåer." Men studien viste bare hvordan endringer i de cis-regulerende elementene i fruktflue-DNA kan ha påvirket fargen på vingestedene i flere forskjellige typer flygende insekter. Den rapporterte ingen vesentlig endring i form eller kroppsplan for disse insektene. I stedet fremhevet studien et tydelig tilfelle av en levedyktig mutasjon som bare genererer en mindre eller overfladisk endring.
Ikke overraskende erkjenner mange evolusjonsbiologer at slike regulatoriske mutasjoner ikke forklarer utviklingen av nye kroppsplaner. For eksempel har Hopi Hoekstra, fra Harvard University, og Jerry Coyne, to tradisjonelle neo-darwinister, publisert en artikkel som gjennomgår forskjellige Evo-Devo-forslag i tidsskriftet Evolution. De bemerker, "Genomiske studier gir liten støtte til den cis-regulerende teorien" om evolusjonsendring.
De argumenterer også, typisk, at de fleste cis-regulerende mutasjoner resulterer i tap av genetiske og anatomiske egenskaper, inkludert et kjent tilfelle der evolusjonsbiologer tilskrev tapet av bekkenrygg i stickleback-fisk til mutasjoner i cis-regulerende elementer. Likevel, som de hevder, "å støtte Evo-Devo hevder at CIS-regulerende endringer er ansvarlige for morfologiske innovasjoner, krever å vise at promotører er viktige i utviklingen av nye egenskaper, ikke bare i tapene av gamle." Hoekstra og Coyne konkluderer med, "Det er for tiden ingen bevis for at cis-regulerende endringer spiller en viktig rolle, -mye mindre en fremtredende en-i adaptiv evolusjon." Gitt deres engasjement for neo-darwinisme, er det rettferdig å anta at Hoekstra og Coyne sannsynligvis ikke hadde til hensikt å komme med dette argumentet, å tilbakevise NCSEs kritikk av vår lærebok Explore Evolution. Likevel sørger vitenskap, likesom politikk, noen ganger for rare 'sengekamerater'.
Bilde 3. Ulike typer regulering styrer cellenes utvikling
Hva med Hox -gener?
Når biologistudenter hører min kollega Paul Nelson beskriver det "store darwinistiske paradokset" på offentlige forelesninger på universitetscampusene, spør de ofte: "Hva med Hox -gener?" Husk at Hox (eller homeotiske) gener regulerer ekspresjonen av andre proteinkodende gener under prosessen med dyreutvikling. Noen biologer har sammenlignet dem med dirigenten til et orkester som spiller rollen i å koordinere musikernes bidrag. Og fordi Hox-gener påvirker så mange andre gener, tror mange Evo-Devo talsmenn at mutasjoner i disse genene kan generere storskala endringer i form.
For eksempel påkaller Jeffrey Schwartz ved University of Pittsburgh mutasjoner i Hox -gener for å forklare det plutselige oppdukkingen til dyreformer i fossilregisteret. I sin bok Sudden Origins erkjenner Schwartz diskontinuitetene i fossilregisteret. Som han bemerker, "Vi er fremdeles i mørket om opprinnelsen til de fleste hovedgrupper av organismer. De vises i fossilregisteret slik Athena gjorde fra hodet til Zeus-fullt ferdig og klar til å gå, i strid med Darwins skildring av evolusjonen som et resultat av den gradvise akkumuleringen av utallige uendelig små varianter. "
Hva løser dette mysteriet? Schwartz, en Evo-Devo-talsmann, avslører svaret: "En mutasjon som påvirker aktiviteten til et homeobox [Hox] -gen kan ha en dyp effekt-for eksempel å vri ... larve-lunikater til de første chordater (tunikater). Det er klart at de potensielle hox-genene har for å vedta det vi kaller evolusjonsendring, ser ut til å være nesten ufattelig. "
Men kan mutasjoner i Hox -gener forvandle en form for dyreliv - en kroppsplan - til en annen? Det er flere grunner til å tvile på at de kan.
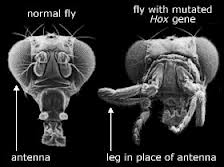 For det første, nettopp fordi Hox -gener koordinerer ekspresjonen av så mange andre forskjellige gener, har eksperimentelt genererte mutasjoner i HOX -gener vist seg å være skadelige. William McGinnis og Michael Kuziora, to biologer som har studert effekten av mutasjoner på Hox -gener, har observert at i fruktfluer "forårsaker de fleste mutasjoner i homeotiske [Hox] -gener dødelige fødselsdefekter." I andre tilfeller er den resulterende Hox -mutante fenotypen, selv om den er levedyktig på kort sikt, likevel markant mindre passform enn den ville typen. For eksempel, ved å mutere et Hox -gen i en fruktflue, har biologer produsert den dramatiske antennapedia -mutanten, en ulykkelig flue med ben som vokser ut av hodet der antennene skal være. Andre Hox -mutasjoner har produsert fruktfluer der balansere (bittesmå strukturer bak vinger som stabiliserer insektet i flyging, kalt "haltere") blir forvandlet til et ekstra par vinger. Slike mutasjoner endrer strukturen til dyret, men ikke på en gunstig eller permanent arvelig måte. Antennapedia -mutanten kan ikke overleve i naturen; Det har problemer med å reprodusere, og avkommet dør lett. Tilsvarende mangler fruktflysmutanter som har et ekstra sett med vinger muskulaturen for å benytte seg av dem, og uten balanse kan den ikke fly. Som den ungarske evolusjonsbiologen Eörs Szathmáry bemerker med forsiktig underdrivelse i tidsskriftet Nature, "Makromutasjoner av denne typen [dvs. i Hox -gener] er sannsynligvis ofte maladaptive."
For det første, nettopp fordi Hox -gener koordinerer ekspresjonen av så mange andre forskjellige gener, har eksperimentelt genererte mutasjoner i HOX -gener vist seg å være skadelige. William McGinnis og Michael Kuziora, to biologer som har studert effekten av mutasjoner på Hox -gener, har observert at i fruktfluer "forårsaker de fleste mutasjoner i homeotiske [Hox] -gener dødelige fødselsdefekter." I andre tilfeller er den resulterende Hox -mutante fenotypen, selv om den er levedyktig på kort sikt, likevel markant mindre passform enn den ville typen. For eksempel, ved å mutere et Hox -gen i en fruktflue, har biologer produsert den dramatiske antennapedia -mutanten, en ulykkelig flue med ben som vokser ut av hodet der antennene skal være. Andre Hox -mutasjoner har produsert fruktfluer der balansere (bittesmå strukturer bak vinger som stabiliserer insektet i flyging, kalt "haltere") blir forvandlet til et ekstra par vinger. Slike mutasjoner endrer strukturen til dyret, men ikke på en gunstig eller permanent arvelig måte. Antennapedia -mutanten kan ikke overleve i naturen; Det har problemer med å reprodusere, og avkommet dør lett. Tilsvarende mangler fruktflysmutanter som har et ekstra sett med vinger muskulaturen for å benytte seg av dem, og uten balanse kan den ikke fly. Som den ungarske evolusjonsbiologen Eörs Szathmáry bemerker med forsiktig underdrivelse i tidsskriftet Nature, "Makromutasjoner av denne typen [dvs. i Hox -gener] er sannsynligvis ofte maladaptive."
Bilde 4. Eks. på maladaptiv hox-mutasjon
For det andre uttrykkes Hox -gener i alle dyreformer etter begynnelsen av dyreutviklingen, og godt etter at kroppsplanen har begynt å bli etablert. I fruktfluer, når Hox -gener kommer til uttrykk, har omtrent 6000 celler allerede dannet seg, og den grunnleggende geometrien til flua - dens fremre, bakre, rygg og ventrale akser - er allerede godt etablert. Så Hox-gener bestemmer ikke dannelse av kroppsplan. Eric Davidson og Douglas Erwin har påpekt at HOX-genuttrykk, selv om det er nødvendig for riktig regional eller lokal differensiering i en kroppsplan, forekommer mye senere under embryogenese enn global kroppsplanspesifikasjon i seg selv, som er regulert av helt forskjellige gener. Dermed er den primære opprinnelsen til dyrekroppsplaner i den kambriske eksplosjonen ikke bare et spørsmål om Hox -genvirkning, men av utseendet til mye dypere kontrollelementer - Davidsons "utviklingsgenerreguleringsnettverk" (Dgrn). Og likevel, som vi så i kapittel 13, hevder Davidson at det er ekstremt vanskelig å endre DGRN -er uten å skade deres evne til å regulere dyreutviklingen. Darwins tvil, s. 312-320;
Stephen Meyer om DGRNS
På dette tidspunktet er det verdt å fortelle hva Meyer skriver om DGRN i kapittel 13 i Darwins Doubt:
Utviklings-generreguleringsnettverk
 En annen forskningslinje i utviklingsbiologi har avslørt en relatert utfordring til den kreative kraften i den neo-darwinistiske mekanismen. Utviklingsbiologer har oppdaget at mange genprodukter (proteiner og RNA) som er nødvendige for utvikling av spesifikke dyrekroppsplaner overfører signaler som påvirker måten individuelle celler utvikler og differensierer seg. I tillegg påvirker disse signalene hvordan celler er organisert og samhandler med hverandre under embryologisk utvikling. Disse signalmolekylene påvirker hverandre til å danne kretsløp eller nettverk av koordinert interaksjon, omtrent som integrerte kretsløp på et kretskort. For eksempel, nøyaktig når et signalmolekyl blir overført, avhenger ofte av når et signal fra et annet molekyl mottas, noe som igjen påvirker overføringen av fortsatt andre-som alle er koordinert og integrert for å utføre spesifikke tidskritiske funksjoner. Koordinering og integrering av disse signalmolekylene i celler sikrer riktig differensiering og organisering av distinkte celletyper under utviklingen av en dyrekroppsplan.
En annen forskningslinje i utviklingsbiologi har avslørt en relatert utfordring til den kreative kraften i den neo-darwinistiske mekanismen. Utviklingsbiologer har oppdaget at mange genprodukter (proteiner og RNA) som er nødvendige for utvikling av spesifikke dyrekroppsplaner overfører signaler som påvirker måten individuelle celler utvikler og differensierer seg. I tillegg påvirker disse signalene hvordan celler er organisert og samhandler med hverandre under embryologisk utvikling. Disse signalmolekylene påvirker hverandre til å danne kretsløp eller nettverk av koordinert interaksjon, omtrent som integrerte kretsløp på et kretskort. For eksempel, nøyaktig når et signalmolekyl blir overført, avhenger ofte av når et signal fra et annet molekyl mottas, noe som igjen påvirker overføringen av fortsatt andre-som alle er koordinert og integrert for å utføre spesifikke tidskritiske funksjoner. Koordinering og integrering av disse signalmolekylene i celler sikrer riktig differensiering og organisering av distinkte celletyper under utviklingen av en dyrekroppsplan.
Følgelig, akkurat som mutering av et individuelt regulatorisk gen tidlig i utviklingen av et dyr, uunngåelig vil stenge utviklingen, vil også mutasjoner eller endringer i hele nettverket av samspillende signalmolekyler ødelegge et utviklende embryo.
Ingen biolog har undersøkt den regulatoriske logikken i dyreutvikling dypere enn Eric Davidson, ved California Institute of Technology. Tidlig i karrieren, og samarbeidet med molekylærbiolog Roy Britten, formulerte Davidson en teori om "genregulering for høyere celler." Med "høyere celler" mente Davidson og Britten de differensierte, eller spesialiserte celler som finnes i ethvert dyr etter de tidligste stadiene av embryologisk utvikling. Davidson observerte at cellene til et individuelt dyr, uansett hvor variert i form eller funksjon, "inneholder generelt identiske genomer." I løpet av en organisme for livssyklus uttrykker genomene til disse spesialiserte cellene bare en liten brøkdel av deres DNA til enhver tid og produserer forskjellige RNA -er som et resultat. Disse fakta antyder sterkt at et dyromfattende system med genetisk kontrollfunksjoner for å slå spesifikke gener av og på etter behov gjennom hele organismens levetid-og at et slikt system fungerer under utviklingen av et dyr fra egg til voksen ettersom forskjellige celletyper blir konstruert.
Bilde 5. Regulering av gener -foregår i genom-nettverk
Da de foreslo sin teori i 1969, erkjente Britten og Davidson at "lite er kjent. . . av de molekylære mekanismene som genuttrykk kontrolleres i differensierte celler. " Likevel utledet de at et slikt system må være på jobb. Gitt: (1) at titalls eller hundrevis av spesialiserte celletyper oppstår under utviklingen av dyr, og (2) at hver celle inneholder samme genom, resonnerte de (3) at en type kontrollsystem må bestemme hvilke gener som er uttrykt i forskjellige celler På forskjellige tidspunkter for å sikre differensiering av forskjellige celletyper fra hverandre-må en eller annen systemomfattende reguleringslogikk føre tilsyn med og koordinere uttrykket av genomet.
Davidson har viet sin karriere til å oppdage og beskrive mekanismene som disse systemene for genregulering og kontrollarbeid under embryologisk utvikling. I løpet av de to siste tiårene har forskning innen genomikk avdekket at ikke-proteinkodende regioner av genomkontrollen og regulerer tidspunktet for uttrykket av proteinkodende regioner i genomet. Davidson har vist at de ikke-proteinkodende regionene av DNA som regulerer og kontrollerer genuttrykk og de proteinkodende regionene i genomet, sammen fungerer som kretsløp. Disse kretsløpene, som Davidson kaller "utviklingsgenerreguleringsnettverk" (eller DGRN) kontrollerer den embryologiske utviklingen av dyr.
Da han ankom Caltech i 1971, valgte Davidson Purple Sea Urchin, Strongylocentrotus purpuratus, som hans eksperimentelle modellsystem. Biologien til S. purpuratus gjør det til et attraktivt laboratoriefag: arten forekommer rikelig langs stillehavskysten, produserer enorme mengder lett befruktede egg i laboratoriet og lever i mange år. Davidson og hans kolleger var banebrytende for teknologien og eksperimentelle protokoller som kreves for å dissekere kråkebollenes genetiske reguleringssystem.
Bilde 6. Dgrn (Development Gen Regulatury Network)
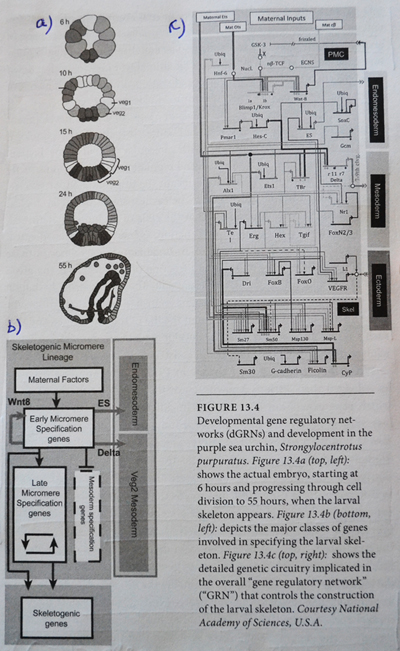 Den bemerkelsesverdige kompleksiteten til det de fant må avbildet visuelt. Figur 13.4a viser urchinembryoet som det ser ut til seks timer etter at utviklingen har startet (øverst til venstre for diagram). Dette er 16-cellestadiet, noe som betyr at fire runder med celledeling allerede har skjedd (1 → 2 → 4 → 8 → 16). Når utviklingen fortsetter i de neste fire trinnene, øker både antall celler og graden av cellulær spesialisering, inntil 55 timer, kommer elementer i urchinskjelettet i fokus. Figur 13.4b viser, tilsvarende disse tegningene av embryoutvikling, et skjematisk diagram med hovedklassene av gener (for celle- og vevstyper) representert som bokser, koblet med kontrollpiler. Til slutt viser figur 13.4c hva Davidson kaller "den genetiske kretsløpet" som slår på de spesifikke biomineralisering-genene som produserer de strukturelle proteinene som er nødvendige for å bygge urchinskjelettet.
Den bemerkelsesverdige kompleksiteten til det de fant må avbildet visuelt. Figur 13.4a viser urchinembryoet som det ser ut til seks timer etter at utviklingen har startet (øverst til venstre for diagram). Dette er 16-cellestadiet, noe som betyr at fire runder med celledeling allerede har skjedd (1 → 2 → 4 → 8 → 16). Når utviklingen fortsetter i de neste fire trinnene, øker både antall celler og graden av cellulær spesialisering, inntil 55 timer, kommer elementer i urchinskjelettet i fokus. Figur 13.4b viser, tilsvarende disse tegningene av embryoutvikling, et skjematisk diagram med hovedklassene av gener (for celle- og vevstyper) representert som bokser, koblet med kontrollpiler. Til slutt viser figur 13.4c hva Davidson kaller "den genetiske kretsløpet" som slår på de spesifikke biomineralisering-genene som produserer de strukturelle proteinene som er nødvendige for å bygge urchinskjelettet.
Dette siste diagrammet representerer et utviklingsgenettverk (eller DGRN), et integrert nettverk av protein og RNA-signaleringsmolekyler som er ansvarlige for differensiering og arrangement av de spesialiserte cellene som etablerer det stive skjelettet til kråkebollen. Legg merke til at for å uttrykke biomineraliserings-genene som produserer strukturelle proteiner som utgjør skjelettet, må gener langt oppstrøms, aktivert i utvikling mange timer tidligere, først spille sin rolle.
Denne prosessen skjer heldigvis ikke tilfeldigvis i kråkeboller, men via sterkt regulerte og presise kontrollsystemer, som det gjør hos alle dyr. Selv et av de enkleste dyrene, ormen C. elegans, som har drøyt 1000 celler som voksen, er konstruert under utvikling av DGRN-er av bemerkelsesverdig presisjon og kompleksitet. Hos alle dyr dirigerer de forskjellige DGRN -ene det Davidson beskriver som embryoets "progressive økning i kompleksitet" - en økning, skriver han, som kan måles i "informasjonsmessige termer."
Davidson bemerker at kompleksiteten til DGRN -ene som er etablert, gjør dem hardnakket motstandsdyktige mot mutasjonsendring - et punkt han har understreket i nesten alle publikasjoner om emnet de siste femten årene. "I kråkebolle-embryoet," påpeker han, "gir det å avvæpne en av disse underkretsene en viss abnormitet i uttrykket."
Utviklingsgenerreguleringsnettverk motstår mutasjonsendring fordi de er hierarkisk organisert. Dette betyr at noen utviklingsmessige gen -regulatoriske nettverk kontrollerer andre genregulerende nettverk, mens noen bare påvirker de individuelle genene og proteiner under deres kontroll. I sentrum av dette regulatoriske hierarkiet er regulatoriske nettverk som spesifiserer aksen og den globale formen for dyre kroppsplanen under utvikling. Disse DGRN -ene kan ikke variere uten å forårsake katastrofale effekter på organismen.
Det er faktisk ingen eksempler på disse dypt forankrede, funksjonelt kritiske kretsløpene som i det hele tatt varierer. I periferien av hierarkiet er genreguleringsnettverk som spesifiserer ordningene for mindre funksjoner som noen ganger kan variere. Likevel krever det å produsere en ny kroppsplan å endre dyrens akse og global form. Dette krever å mutere selve kretsløpene som ikke varierer uten katastrofale effekter. Som Davidson understreker, fører mutasjoner som påvirker DGRN-ene som regulerer utvikling av kroppsplan til "katastrofalt tap av kroppsdeler eller fullstendig tap av levedyktighet." Han forklarer mer detaljert:
"Det er alltid en observerbar konsekvens hvis en DGRN -underkrets blir avkuttet. Siden disse konsekvensene alltid er katastrofalt dårlige, er fleksibiliteten minimal, og siden underkretsene er sammenkoblet, tar hele nettverket i kvaliteten på at det bare er en måte for ting å fungere på. Og faktisk utvikler embryoene til hver art seg på bare en måte."
Ingeniørbegrensninger
Davidsons funn gir en dyp utfordring for den neo-darwinske mekanismen. å bygge en ny dyrekroppsplan krever ikke bare nye gener og proteiner, men nye DGRN -er. Men for å bygge en ny DGRN fra en eksisterende Existing DGRN ved mutasjon og seleksjon krever nødvendigvis å endre det eksisterende utviklingsgenetten nettverket (selve typen endring som vi så i kapittel 12 ikke kan oppstå uten flere koordinerte mutasjoner). I alle fall har Davidsons arbeid også vist at slike endringer uunngåelig har katastrofale konsekvenser.
Davidsons arbeid fremhever en dyp motsetning mellom den neo-darwinistiske beretningen om hvordan nye dyrekroppsplaner er bygget og et av de mest grunnleggende prinsippene for ingeniørfag-prinsippet om begrensninger. Ingeniører har lenge forstått at det mer funksjonelt integrerte systemet er, jo vanskeligere er det å endre noen del av det uten å skade eller ødelegge systemet som helhet. Davidsons arbeid bekrefter at dette prinsippet gjelder for å  utvikle organismer .. Systemet med genregulering som kontrollerer utvikling av dyre-kroppsplaner er utsøkt integrert, slik at betydelige endringer i disse genregulerende nettverkene uunngåelig skader eller ødelegger det utviklende dyret. Men gitt dette, hvordan kan en ny dyrekroppsplan, og de nye DGRN -ene som er nødvendige for å produsere den, noen gang utvikle seg gradvis via mutasjon og seleksjon fra en eksisterende kroppsplan og sett med DGRN -er?
utvikle organismer .. Systemet med genregulering som kontrollerer utvikling av dyre-kroppsplaner er utsøkt integrert, slik at betydelige endringer i disse genregulerende nettverkene uunngåelig skader eller ødelegger det utviklende dyret. Men gitt dette, hvordan kan en ny dyrekroppsplan, og de nye DGRN -ene som er nødvendige for å produsere den, noen gang utvikle seg gradvis via mutasjon og seleksjon fra en eksisterende kroppsplan og sett med DGRN -er?
Davidson gjør det klart at ingen egentlig vet: "I motsetning til klassisk evolusjonsteori, kan ikke prosessene som driver de små endringene som er observert i det arter divergerer tas som modell for utviklingen av kroppsplanene til dyr."
Bilde 7. Davidson var kritisk til gen-mutasjoner som årsak til formednring
Han utdyper:
"Neo-Darwinistisk Evolusjon. . . Antar at alle prosesser fungerer på samme måte, slik at utviklingen av enzymer eller blomsterfarger kan brukes som nåværende fullmakt for å studere utviklingen av kroppsplanen. Det antar feilaktig at endring i proteinkodende sekvens er den grunnleggende årsaken til endring i utviklingsprogram; og det antar feilaktig at evolusjonsendring i kroppsplan-morfologi skjer ved en kontinuerlig prosess. Alle disse forutsetningene er i utgangspunktet kontrafaktiske. Dette kan ikke være overraskende, siden den neo-darwinistiske syntesen som disse ideene stammer fra, var en premolekylær biologi, fokusert på populasjonsgenetikk og ... naturhistorie, som ingen av dem har noen direkte mekanistisk import for de genomiske reguleringssystemene som driver embryonisk utvikling av kroppsplanen."
Darwins Doubt, s. 264-269
 Casey Luskin (Bilde 8).
Casey Luskin (Bilde 8).
Casey Luskin er geolog og advokat med høyere grad i vitenskap og jus, som gir ham ekspertise i både de vitenskapelige og juridiske dimensjonene av debatten om evolusjon. Han fikk sin doktorgrad i geologi fra University of Johannesburg, og BS- og MS-grader i geovitenskap fra University of California, San Diego, hvor han utstrakt studerte evolusjon, både på hoved- og lavere nivå. Hans jusgrad er fra University of San Diego, hvor han fokuserte studiene på 1.Amendment, utdanningslov og miljørett.
Oversettelse, via google oversetter, og bilder ved Asbjørn E. Lund