Et tilsvar til Hans Øvland Tjelle
Først får vi si at Tjelle kommer med mye interessant informasjon, som ikke nødvendigvis er kjent for alle. Det blir et sammenstøt mellom to ulike ulike teorier, med hver sine representanter. Om representanten for evolusjonsteorien synes best orientert, betyr det ikke nødvendigvis at teorien han forfekter er den riktige. Ikke heller at forskjellen mellom målbærerne for teoriene er så stor som han synes å påstå.
 1. Morfologi:
1. Morfologi:
Det finnes andre forklaringer enn nedarving ved tilpasning på morfologiske likheter (homologi). Før Darwin så en gjerne at samme design var benyttet i ulike organismer. En enkel oppstilling av former som ligner hverandre, gir ikke en forklaring på opphavet i seg selv. Det kreves en mekanisme som kan forklare slike likheter. Forklaringer som "nedstamning og tilpasning" er bare ord, om de ikke kan kobles mot faktiske biologiske prosesser. De fleste biologer før Darwin ville ha forklart slike likheter med felles design eller skapelse. Fossilmaterialet er forenlig med begge forklaringer.
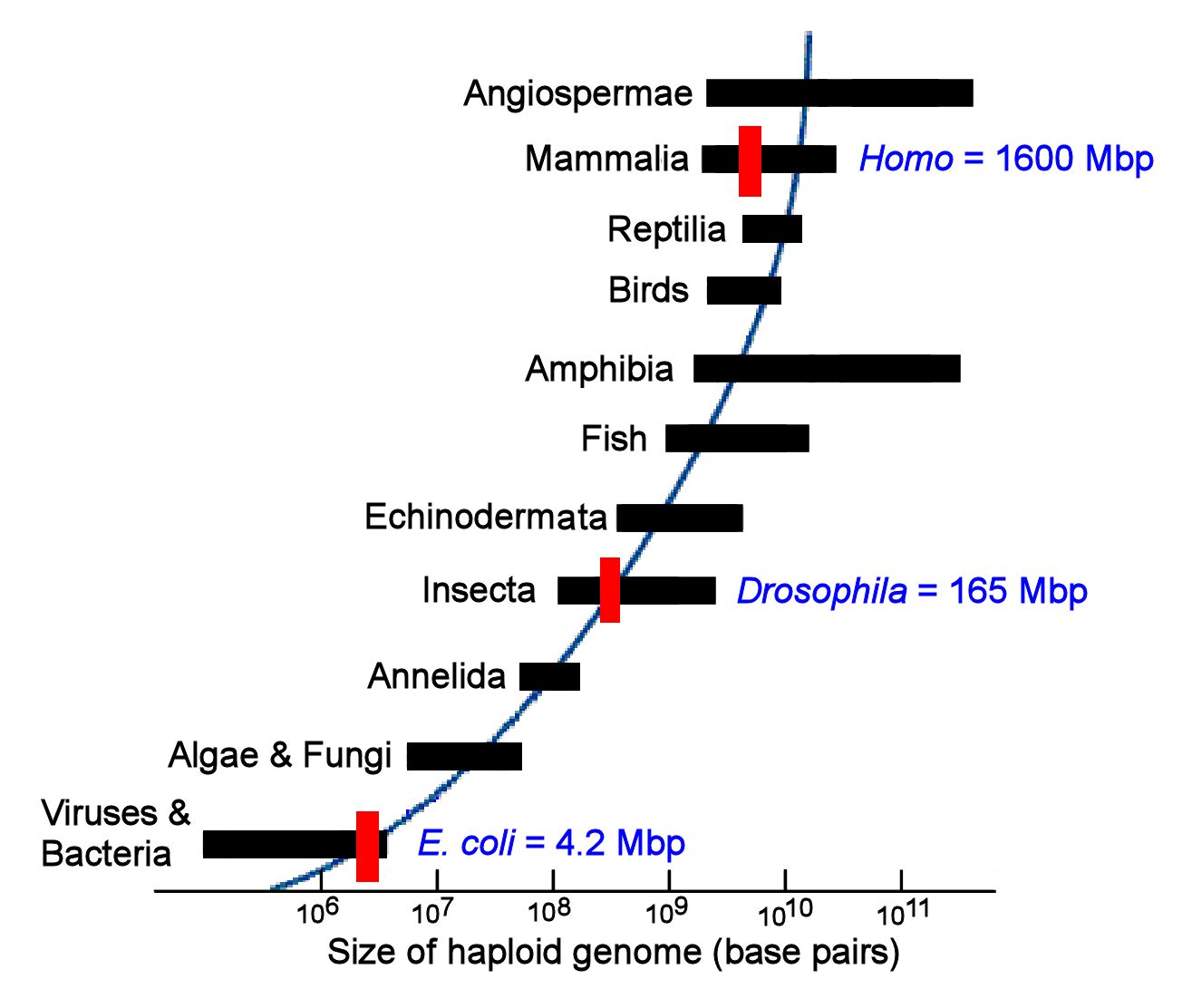
Bilde 1. Variabel størrelse på genomet
C-verdi paradokset
'Massen av DNA i en organismes (haploide) kjønnscelle; organismens "C-verdi'
C-verdien er vanligvis uttrykt i picogram (pg), som er en billionte-del av et gram. Det er 978 millioner base-par i et picogram. C-verdien til mennesker er ca. 3.5 pg, som tilsvarer ca. 3.2 milliarder basepar. DNA i cellene til en organisme, blir vanligvis forstått å inneholde informasjon nødvendig for dens utvikling og funksjon. Det synes derfor fornuftig å anta at mengden til en organismes DNA, er et slags mål på dens kompleksitet. Men det synes ikke å være et konsistent forhold mellom C-verdien og den tilsynelatende kompleksiteten til organismen. Denne inkonsistensen har vært betraktet som et paradoks. Det syntes problematisk at C-verdien for noen dyr var betydelig større enn for mennesket. Verdien for den blå-flekkede salamanderen (Ambystoma laterale), er 81 pg, eller 23 ganger større enn for mennesket. C-verdi for amerikansk hulesalamander (Necturus lewisi) er 120 pg, eller 34 ganger større enn for mennesket. Det er ingen grunn til å anta at utvikling og virkemåte til disse dyrene, f.eks. større tilpasningsdyktighet i forhold til mijøendringer, er mer komplekse enn for et menneske. Det synes dermed merkelig at disse dyrene skulle ha så mye mer DNA enn ett menneske. Denne gåten, kjent som C-verdi paradokset , stammer fra Thomas (1971).
Like merkelig er det også at selv noen 'nære slektninger' til dyr eller planter har svært annerledes C-verdier. F.eks. de to paddene (Scaphiopus couchii) og den europeiske (Bombina bombina). Her har den første har en C-verdi på nær 1, mens den andre har en C-verdi på mer enn 12. Likedan for den utsmykkede hule-frosken (Ceratophrys ornata), den første har en C-verdi på omkring 1, mens den horn-utsmykket slektning (Limnodynastes ornatus) har C-verdi på mer enn 13. Det er mange andre eks. på parallelle arter med lignende morfologi, som har vidt ulike C-verdier. Variasjonen i C-verdi over hele klassen amfibier har en faktor på 120. (-fra 'Animal Genome Database: genomesize.com). Hvorfor skulle meget like dyrearter, ha slike ulike C-verdier? Om én av dem kan utvikle seg og fungere med mindre DNA-innehold, hvorfor trenger den andre ha så mye mer?

Bilde 2 og 3. To frosker med kvotient 13 i C-verdi

Konvensjonell visdom' er at ulikhet i C-verdier kan tilskrives store mengder av ubrukelig eller 'søppel-DNA' i noen arter og mindre i andre.. Men disse svarene kommer til kort, i forhold til ENCODE-programmet, som rapporterte at 'søppel-DNA' ikke er søppel. Darwinisters argument er sirkulært: eksistensen av 'søppel-DNA' sies å løse C-verdi paradokset, mens C-verdi paradokset blir brukt som et argument for validiteten av 'søppel-DNA'. Men et sirkelargument er ingen løsning. For darwinister må C-verdi problemet være et paradoks.
Endelig kan vi vel være enige om at kua, morfologisk sett, har større likheter med hesten enn med hvalen.
2.A Genetikk
For Cytokrom C sitt vedkommende måtte jeg (som ikke-biolog), søke opp forklaring på dette, som jeg best fant hos creation.com. Jeg fant ikke nå, det jeg fant da, som kan ha vært noe villedende, men det jeg fant var en interessant kobling opp mot Haldane's dilemma, som med realistiske rater begrenser antallet tjenlige mutasjoner som det kan gjøres rede for oppad til en grense på 1.667 mutasjoner for menneskelig evolusjon, som kan ha inntruffet på de antatt 10 millioner år siden 'siste felles stamfar' mellom mennesker og aper.
Det innebærer bare en substitusjon pr. 300 generasjoner i gjennomsnitt. Opprinnelsen til alt som gjør oss unikt menneskelige må forklares innen den grensen. Biologer har funnet at den store majoritet av substitusjoner er punkt-mutasjoner, så Haldanes grense setter en alvorlig begrensning på hva som er mulig med evolusjon, fordi 1.667 enkelt mutasjoner utgjør mindre enn et gen av gjennomsnittlig størrelse.
Se mer om Haldanes dilemma -her.
Enn videre: Forskning viser at alle arter har gener, micro-RNA og proteiner som er spesifikke for arten. De finnes altså ikke andre steder i dyreriket. De er unike, og kan bety tilførsel av ny spesifikk informasjon til organismer. De ca. 25-30.000 gener hos pattedyr kan kode for over en million ulike protein-varianter. Det skyldes den enorme kompleksitet og oppfinnsomhet til stede i prosesser, i form av f.eks. alternativ transkripsjon og spleising av RNA avskrifter/ kopier.
Tidligere anså man ‘ikke-protein kodende’ regioner av genomet som ubrukelige. Men nå er det kjent de utfører en rekke helt nødvendige oppgaver. Micro-RNA, som består av ca. 22 RNA-bokstaver, kan spille stor rolle for genfunksjon. Singelton proteiner, som kun finnes i en art, og ikke hos andre arter, er forenlig med at hver art er unik.
Disse funksjonene går på: 1. Regulering av DNA-replikasjon (27). 2) Regulering av transkripsjon/-koding (28) 3) Markering av lokasjoner for programmerte rearrangement av genetisk materiale (29). 4) Påvirke helt nødvendig tre-dimensjonal folding og vedlikehold av kromosomer (30) 5) Kontrollere interaksjon mellom kromosomer og kjernemembran og matrix (31). 6) kontrollere RNA prosessering -editering og spleising (32) 7) Modulering: I cellen dreier det seg om oversetting via m-DNA og t-DNA i forhold til RNA (33). 8) Regulering av embryo-utvikling (34) 9) Reparering av DNA (35) 10) Hjelp til immunforsvar eller sykdomsbekjempelse (36). Det ikke-protein kodende del av genomet fungerer mye likt et operativsystem i en PC og kan styre flere operasjoner simultant (37).
Californske vitenskapsmenn undersøkte i 2007: 10.402 ikke-kodende elementer av det menneskelige genom og fant at en forbausende høy prosentandel fungerte i forbindelse med regulering av gener. De konkluderte med at mobile elementer kan ha spilt en større rolle enn tidligere gjenkjent. (14) det samme året kunne australske molekylær-biologer rapportere: "Mens mindre enn 1,5% av genomet koder for protein, er det nå klart at den store majoriteten blir transkribert, hovedsakelig til ikke-protein kodende RNA hvorav et stigende antall viser seg å være funksjonelle. (15) Og i 2008 demonstrerte amerikanske forskere en viktig funksjon for ikke-kodende RNA kopiert fra repeterende segmenter av DNA, som tidligere var betraktet som søppel-DNA. (16)
Bilde 4. Ikke-protein kodende del av genom kan si vel så mye om organismen.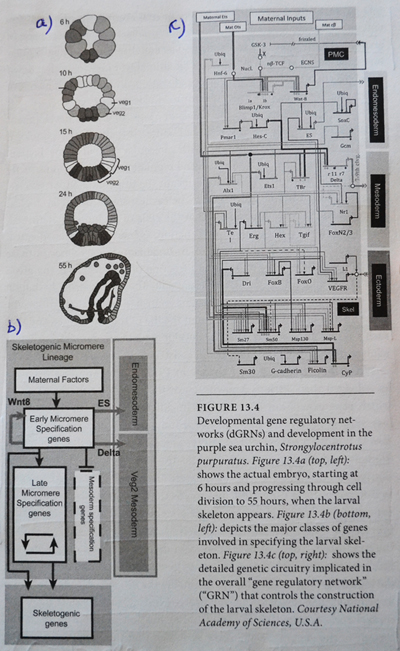
Gjennom de siste tiårene har forskning i genomet avslørt at ‘ikke-kodende’ regioner av genomet, kontrollerer og regulerer timingen til hvordan protein-kodende regioner av genomet uttrykker seg. Sammen fungerer ikke-protein kodende og protein-kodende regioner av genomet som et nettverk i følge Davidson. Davidson valgte fra 1971 et sjøpiggsvin (Strongylocentrotus purpuratus) som modell for sitt eksperiment. Sammen med kolleger utviklet Davidson et pioner-prosjekt og eksperimentelle protokoller som krevdes for å dissekere og kartlegge sjøpiggsvinets genetiske regulerende nettverk. Den fantastiske kompleksiteten de fant framstilles på figure 13.4 under til høyre. (Dgrns)
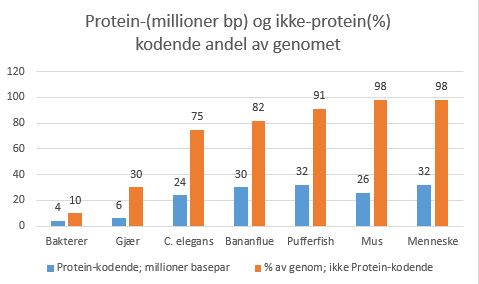
Noe av det viktigste som ny forskning innen epigenetikk bringer oss, er alle de måter gener kan uttrykkes på. Da er det ikke lenger snakk om tilfeldige mutasjoner, som for alle praktiske forhold har vist å bringe uorden, sykdom og død. Nei, det dreier seg f.eks. om at det er mange måter å kombinere oppskriften i DNAet på. En sammenligner med script som er tilpasset de spesifikke celletyper, over 550 i mennesket, og at RNA har en fundamentalt større betydning enn man tidliger har tillagt det. Tidligere ble RNA betraktet som midlertidige 'budbærere' (mRNA) og 'transportører' (tRNA), hvor alt dreide seg om DNA-etsproduksjon av proteiner. Bananflue har ca. 30 millioner proteinkodende basebar, mens mus bare har 26 mill. og mennesket 32 millioner. Bakterier har ca. 10% av genomet som ikke koder for proteiner, gjær ca. 30%, nevnte C. elegans har 75%, bananflue har ca. 82, 'pufferfish' har 91, mus og menneske har ca. 98%. Vi ser at økende kompleksitet er mer i samsvar med andel av genomet som ikke koder for proteiner, enn det er med antall basepar som koder for proteiner. Det er på høy tid å rette opp i en språklig utydelighet: ikke-kodende DNA innebærer at den ikke koder FOR PROTEIN, ikke nødvendigvis at den ikke koder for noe som helst. Fravær av bevis, er ikke det samme som bevis for fravær. En må ha rett teknologi for å oppdage hva som evt. kodes. Det var ikke før ny teknologi for sporing, kombinert med enormt øket datakraft, at vi begynte å innse at noe meget interessant skjedde i de resterende 98%, den 'ikke-kodende' del av genomet.
En temmelig fersk artikkel fra PLoS Genetics rapporterte om " 60 nye protein-kodende gener som oppsto 'de novo' på den menneskelige linjen, siden atskillelse med sjimpanser". Det var et funn som ble kalt mye større enn ett tidligere, konservativt estimat". Etter en studie av bananfluer som rapporterte at 12% av nylig oppdukkede gener i Drosophila melangaster undergruppen, kan ha oppstått 'de novo' fra ikke-protein kodende DNA." (29) Forfatteren fortsatte her med å erkjenne at å kalle på denne mekanismen stilte ett alvorlig problem for evolusjonsteori, siden det ikke forklarte opprinnelsen til noen av dets 'ikke-trivielle krav for funksjonalitet."
Bilde 5. dGRN's hos sjøpiggsvinet
Et annet eks: nå er evolusjonister møtt med en ny utfordring. Forfatterne av PNAS- papiret, Hughes og Chevance, beskriver hva som drev dem til å undersøke sammenhengen for hvert triplettkodon. De 'lekte seg med' gener for en del av bakterieflagellen ved navn FlgM da de la merke til noe interessant:
"Endring av kodonen på den ene siden av det defekte kodon resulterte i en 10-gangers økning i FlgM proteinaktivitet. Endring av kodonen på den andre siden resulterte i en 20-gangers reduksjon. Og de to endringene sammen ga en 35-ganger økning. "Vi innså at disse to kodonene, selv om de var adskilt av et kodon, snakket til hverandre ," sier Hughes. "Den effektive koden kan være en triplett av tripletter."
Hvis dette gjelder generelt, så må genetikere, for å forstå den genetiske koden, forstå konteksten til hver triplett. Hvert kodon kan bli påvirket av sine flankerende kodoner, noe som skaper svært forskjellige utfall når det gjelder genaktivitet. Mer her. Det synes som mekanismen med mutasjoner i en én-dimensjonal DNA-streng, er altfor enkel til å forklare endringer i livets komplekse virkelighet.
2.B Epigenetikk. Om en skal vurdere vurdere korrektheten til evolusjonsteorien, kan en ikke bare gå utenom nivået over genetikken: epigenetikken:
 Høyere ordens, strukturell informasjon synes spille en avgjørende rolle for utvikling av organismer. Biologer vet ikke hvor alle denne informasjonen utenom genomet befinner seg, men har lokalisert noe. For eks. vet man at strukturen og lokaliseringen til celle-skjelettet spiller inn på utviklingen av embryoet. I tillegg påvirker lokalisering av spesifikke mål-steder på innsiden av celle-membranen formen på celle-skjelletet og influerer utviklingen av organisme-formen. Selv om disse er laget av proteiner, er det ikke bare proteinet, men plasseringen og den tre-dimensjonale strukturelle informasjonen som bestemmer strukturen på celleskjelettet og lokaliseringen av dets mindre enheter.
Høyere ordens, strukturell informasjon synes spille en avgjørende rolle for utvikling av organismer. Biologer vet ikke hvor alle denne informasjonen utenom genomet befinner seg, men har lokalisert noe. For eks. vet man at strukturen og lokaliseringen til celle-skjelettet spiller inn på utviklingen av embryoet. I tillegg påvirker lokalisering av spesifikke mål-steder på innsiden av celle-membranen formen på celle-skjelletet og influerer utviklingen av organisme-formen. Selv om disse er laget av proteiner, er det ikke bare proteinet, men plasseringen og den tre-dimensjonale strukturelle informasjonen som bestemmer strukturen på celleskjelettet og lokaliseringen av dets mindre enheter.
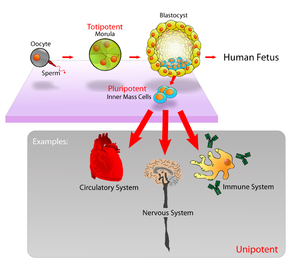
I encellede organismer kan kutt i celle-membranen danne arvelige endringer i membran-mønstre, selv om DNA ikke er skadet. I det hele bestemmes formen og strukturen til cella av både gener og tidligere tredimensjonale-strukturer og deres organisering. Proteiner transporteres og finner fram, til riktig adresse delvis p.g.a. tidligere tredimensjonale-strukturer og deres organisering. Mer her. En har funnet at ca. 50% av lange (ipk)RNA faktisk kunne øke gen-uttrykket for nabogener. Pluripotente celler, som kan utvikle seg til alle typer celler, men unntak av morkaken. Shinya Yamanaka har identifisert gener og proteiner som er uttrykt i ES-celler. Gener som holder cellen i pluripotent tilstand, er meget viktige -fordi ES-celler er meget følsomme. En ørliten endring i betingelsene cellen lever i, og den endrer seg f.eks. til nevroner i hjernen. En annen ørliten endring og de utvikler seg til hjerte-celler.. De identifiserte massevis av celler som var viktige for å holde cellen i pluripotent tilstand. Se evt. mer her.
Bilde 6. Pluripotente celler
Tjelle: "Hva er IDs teori som forklarer divergens over tid i proteinsekvenser kontra divergens over generasjoner i ukodende sekvenser? Hvordan forklarer ID at mønsteret passer overens med morfologiske trær?"
Jeg vet faktisk ikke hvordan ID forklarer dette p.t. Følg med her, så skal jeg dele det om/når det kommer. Når det gjelder 'ukodende sekvenser', henviser jeg til pkt. 2.A ovenfor -'ikke-protein kodende' sekvenser.
3. Fylogenetiske trær
Jeg er glad vi er enige om at: "Det interessante er ikke at fylogenetiske trær er i konflikt med hverandre." Jeg er mer i tvil om metoden å "fjerne det minste mulige antall grener som ikke er kongruente. Ved å gjøre det ender vi opp med å måtte fjerne 4 grener: piggsvin, ekorn, trespissmus og skjermflygere. Resterende grener er en perfekt match." Spillanalogien blir: 'For å få det til å passe, fjerner man det som ikke passer'. Det er sjelden en hører om ekstreme sannsynligheter i positiv valør i forbindlese med evolusjonsteorien. Et mer typisk eks. i forbindelse med sannsynlighet for suksess i evolusjonsteorien er f.eks:
Hvor mange mutanter måtte dukke opp for å sikre deres overlevelse? Dersom vi kjenner deres selektive verdi, kan vi beregne sannsynligheten. En typisk selektiv verdi for den type mutasjoner vi ser på, ble av George Gaylord Simpson, ansett som en evolusjonisthøvding, satt til 0,1% (Simpson 1953, s.119). Som nevnt viste Fishers beregninger at for bare én mutasjon ville sjansen for overlevelse være 1:500. Det måtte være nesten 250 slike for at en mutant skulle ha over 50% sjanse for overlevelse.
 Vi skal repetere hva som kreves for at å fullføre ett av 500 suksessfylte steg: En mutasjon må inntreffe, og den må overleve for å ta over populasjonen. Men sjansen er liten: Sjansen for at den vil inntreffe ble estimert til 1:600. Med en selektiv verdi på 0,1%, så er sjansen for at mutasjonen skal overleve 1:500. Sjansen for at den både skal dukke opp OG overleve er anslagsvis: (1/600)*(1/500) som gir 1:300.00. For at en ny art skal inntreffe, må 500 av disse inntreffe uten å feile. Sjansen for at dette skal skje, blir (1:300.000) opphøyd i 500 som er ca.
Vi skal repetere hva som kreves for at å fullføre ett av 500 suksessfylte steg: En mutasjon må inntreffe, og den må overleve for å ta over populasjonen. Men sjansen er liten: Sjansen for at den vil inntreffe ble estimert til 1:600. Med en selektiv verdi på 0,1%, så er sjansen for at mutasjonen skal overleve 1:500. Sjansen for at den både skal dukke opp OG overleve er anslagsvis: (1/600)*(1/500) som gir 1:300.00. For at en ny art skal inntreffe, må 500 av disse inntreffe uten å feile. Sjansen for at dette skal skje, blir (1:300.000) opphøyd i 500 som er ca. . Minste sannsynlighet for at noe vil inntreffe tilfeldig i virkeligheten, er av W. Dembski satt til
. Minste sannsynlighet for at noe vil inntreffe tilfeldig i virkeligheten, er av W. Dembski satt til 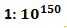 (10 opphøyd i -150). Dette har en sannsynlighet som er av størrelsesorden
(10 opphøyd i -150). Dette har en sannsynlighet som er av størrelsesorden  mindre enn
mindre enn 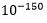 !
!
Ang. hvalers avstamning, må jeg minne om noe jeg nevnte tidligere: I 2007 pekte J.G.M Thewissen og kolleger på at siden hvaler dukker opp i fossil-materialet 35 millioner år tidligere enn flodhesten, er det ikke sannsynlig at de to er nær relatert.
Fig. 1. Hva som med rimelighet kan dannes tilfeldig
Eks. på spesielle forhold ved hval: i) setefødsel -ellers ville ungen druknet (vanskelig 'overgang') ii) hvalen har ikke forbindelse mellom åndedretts-organer og munnhulen: Det gjør at den kan dukke med vidåpen munn, og få i seg masse mat uten å få det i luftrøret.. iii) Hvaler har spesielle organer for dypvanns-dykking: Hvalen klarer å lokalisere byttet nøyaktig ved hjelp av sitt unike sonarsystem. For hvalen kommer 9% av oksygenet fra lungene under dykking, mens 41% kommer fra blodet og 50% fra muskler og vev. (Tilsvarende tall for mennesket: 34% fra lungene, 41% fra blodet og 25% fra muskler/vev). iv) Hvaler har egen øre-konstruksjon: De indre knoklene i øret, har en helt annen form en hos oss. v) Hvaler har en særegen filtrerings-mekanisme: Grønnlandshvalen har ca. 350 barder med lengde opptil 4,5 m. En hval kan høste et tonn krill, fra 10.000 m3 havvann.vi) Noen hvaler kan  komponere musikk.
komponere musikk.
Så selv om Tjelle hevder: "Med andre ord er det stort samsvar mellom DNA og morfologi: alle analyser plasserer hvaler innenfor klovdyr," så oppfordrer jeg til dem som ikke er 'bundet av' evolusjonsteorien, til å vurdere rimeligheten av dette i tråd med informasjonen ovenfor, evt. spe på med mer særtrekk ved hvaler -her.
Om en danner et fylogenetisk tre ut fra proteinet prestin alene, så blir flaggermus og delfiner i slekt. Slike trær korrigeres ut fra evolusjonære forutsetninger, og det er når en kommer til et protein som gir bortimot 'helklaff' (Cytokrom C), så får en høre om det. Siler en så ut 'det som ikke passer', så passer alt -nokså logisk egentlig. Men for alle proteiner gjelder at deres 3-dimensjonale passform, som virkemåten er betinget av, har vist seg svært vanskelig å framkalle ved mutasjoner. Dette ble påvist ved D. Axe sitt forsøk på å mutere en ny virkemåte mellom KbL og BioF proteiner, siden er dette etterprøvd bl. a. av Ann Gauger -her.
Bilde 7. Orfan-gener en avgjørende forskjell
4. Paleontologi
Tjelle: "Det interessante i denne saken her er at sitatet fortsatt trekkes frem, snart 30 år siden Patterson først uttalte seg, uten det etterfølgende sitatet: “The reason is that statements about ancestry and descent are not applicable in the fossil record. Is Archaeopteryx the ancestor of all birds? Perhaps yes, perhaps no: there is no way of answering the question.[12] Det viktige ordet her er “ancestor”. I moderne paleontologi hevder en ikke at en fossil art er stamfaren til en annen. En utelukker det spørsmålet med mindre det er veldig godt grunnlag for å hevde det. Ingen moderne paleontologer vil hevde at archaeopteryx er moderne fuglers direkte stamfar, fordi det spørsmålet ikke engang drøftes."
Det var greit å få med resten av sitatet også, som jeg ikke har hørt referert før, men i stedet for å imøtegå argumentet om manglende stamfedre, blir det begrunnet. Det tjener paleontologien til ære at den ikke uttaler seg uten veldig godt grunnlag. Så er det helt på linje med min forståelse at: "stamfar er ikke det samme som overgangsform". Det er Tjelle som hevder at Archaeopteryx "ikke har noe med dinosaurer å gjøre". Min påstand var: "holder de fleste paleontologer nå for at Archaeopteryx er en unik mellomform, en art av fugler som nå er utdødd." Igjen: det er ikke godt nok grunnlag for å hevde noe om avstamningen, selv om det trekkes frem flere felles kjennetegn. Jeg tviler ikke på at Archaeopteryx står nevnt i flest fyliogenetiske trær i lag med dinosaurene. Spørsmålet er om den med din talemåte: er på en av de 'ikke-kongruente grenene', som burde fjernes fra det fyliogenetiske treet? Når du hevder at dinosaurer har hulrom i knoklene, så er vel dette hulrom etter beinceller og blodårer etc? Levende fugler har jamt over tomrom i knoklene, slik at de har mulighet til å fly.
 Om fugler stammer fra dinosaurer:
Om fugler stammer fra dinosaurer:
"Hans (Barons) videre spekulasjoner på om stegosaurer eller keratopsider kan ha hatt fjær har ingenting med forskningsprosjektet å gjøre." Dette er slutningen fra forfatterens (Barons) egne kommentarer, og gjorde at undertegnede dro det vel langt. Det 'famøse bildet' er byttet med ett annet, som formodentlig er akseptert av flere enn de ivrigste 'dino-fugl' tilhengere. Sjekk selv her.
Bilde 8. Sinosauropteryx-prima -forløper for fugl?
Det kan stemme at i noen tilfelle har evolusjonsteorien kommet med forutsigelser for hvilke funn en vil gjøre hvor. Poenget er hvorvidt vi ville hørt noe til dem, dersom de viste seg ikke å stemme. Ville de blitt publisert under 'eks. på mislykkede søk'? Vi har vel erfaring for at det er de forutsigelser som viser seg å stemme, som får plass -enten det er i media eller i vitenskapelige journaler..
Tjelle: "Så jeg spør, gitt denne rekken av overganger mellom dinosaurer og fugler, hvor kan vi sette en klar grense mellom gruppene? Uansett hvor du setter grensen vil du utelukke nære slektninger som likner sterkt på de dyrene du inkluderer."
Det benektes ikke at det er mange overgangsformer. Som nevnt ovenfor er ikke det nødvendigvis det samme som stamfedre. Om fugler har mest likhet med dinosaurer eller krypdyr, spiller ikke all verdens rolle for Intelligent Design (ID). Fugler kan ha likheter med begge, via et felles design. Dermed ser jeg ikke noe behov for å sette noen grense. Det å sette streker mellom grupper for å knytte dem sammen, og for å skille grupper er noe evolusjonsteorien er gode til -ut fra sine forutsetninger.
Kritikk av påståtte bevis for evolusjonsteorien
Når vi vurderer de ulike funn om kroppsform og utvikling som Tjelle presenterer, er det lite eller ingenting som definitivt støtter trans-spesifikk evolusjon. Likheter eller homologier mellom ulike kroppsformer peker minst like sterkt på en enkelt felles designer, i motsetning til mange designere. Også fellestrekkene ville til og med bringe en slik designer stor ære i de fleste kulturer, noe som indikerer hans mestring av alle sine design via ulike former. Denne vinklingen styrkes når man ser på den transcendentale kompleksiteten til selv den enkleste levende cellen -lenke. På samme måte argumenterer likheten mellom mikrobiologiske prosesser i forskjellige arter like mye - faktisk mer - for deres felles design enn for en felles fysisk anestesi. En sitter igjen med følelsen av at evolutionsmodellen gjennomsyrer Tjelles tenkning, slik at han hopper over de mer åpenbare bevis for allestedsnærværende design.
Bilde 9. Om du er vitenskapsperson eller ikke: vurder selv hva vi ser omkring oss
Det har hendt før at en har funnet eldre fossiler enn en har tekt seg, -lenke. Skulle det dukke opp nyheter som er interessante for Intelligent Design, kan du finne dem her. Det dreier seg om felles avstamning eller ikke her. Det dukker stadig opp nye momenter som motsier dette -lenke. Hvordan vi oppfatter dette, har mye med hva vi har tro for. Jeg har ikke så stor tro på blinde, ikke-styrte naturprosesser som Tjelle synes å ha.
Konklusjon:
Om noen er uenige i evolusjonsteorien, er de nærmest garantert å få høre at de mangler forståelse for hva den er, -samt manglende kunnskap i biologiske vitenskaper. Det synes å komme som en refleks, om en er uenig i standard-teorien. Når det påstås at nye former dukker opp uten kjente forgjengere i Kambrium, blir ikke det bestridt av Tjelle -så vidt jeg ser. Hva kan oppkomsten av disse skyldes? Siden diskusjonen fant sted i apologetisk gruppe: Er det mulig at 'igangsetteren/den første årsak' har 'fulgt opp sitt skaperverk' -i stedet for å overlate det til naturens blinde prosesser? Jeg vil påstå at ID bedre fanger opp dataene vi observerer, f.eks. i forbindelse med såkalt 'junk-DNA', epigenetikk og i alle sammenhenger der evolusjonsteorien sliter: Den har bare ett trick i ermet (naturlig seleksjon), og har få og dårlige forklaringer på hvordan nødvendig ny informasjon dannes i gener.
Jeg er heller ikke biolog eller paleontolog, men vil ikke av den grunn anbefale lesere å oppsøke populærvitenskapelig litteratur, selv om den er skrevet av fagfolk. Grunnen er at de erfaringsmessig bare serverer 'ferdigtygde argumenter' og springer over interne diskusjoner. Vitenskapsfolk med 'ferdige svar' er ikke nødvendigvis de som har rett eller som tjener vitenskapens interesser best.
Stoffutvalg og bilder ved Asbjørn E. Lund