Det essensielle maskineriet for å starte livet
Luca (Last Universal Common Ancestor): Dechiffrering av roten til livets universelle tre
Oversatt herfra -kilder finnes her.
Innenfor det mangfoldige livets rike (biosfæren) på jorden, postulerer det vitenskapelige samfunnet eksistensen av Luca, eller den siste universelle felles stamfar. Denne enheten står ikke som en organisme fossilisert i eldgamle bergarter, men snarere som en teoretisk konstruksjon, utledet av komparativ genomikk og molekylærbiologi. Karakteriseringen av Luca, som representerer en felles avstamning for alle eksisterende livsformer, gir et glimt av tidlige evolusjonsparadigmer. Det er generelt postulert at Luca levde mellom 3,5 til 3,8 milliarder år siden, basert på molekylære klokkeanalyser og eldgamle fossile bevis. Luca markerer imidlertid ikke livets opprinnelse; I stedet betyr det en tilstand av molekylær modenhet, og understreker en mengde biologiske prosesser som allerede var i drift.
 Sammenlignende genomikk gir innsikt i den potensielle genomiske rammen for Luca. Denne stamfaderen ville ha huset gener som er sentralt for RNA -syntese, DNA -replikasjon, proteinproduksjon og rudimentære metabolske prosesser. Slike molekylære mekanismer danner grunnlaget for den livlige trivselen til livet vi observerer i dag. En kjernefunksjon som antas å være til stede i Luca's cellulære maskiner, er dens kapasitet for nukleinsyresyntese og vedlikehold. I motsetning til en forenklet modell, ville det genetiske maskineriet til Luca være kompetent, noe som muliggjør DNA -replikasjon og reparasjon, i likhet med en erfaren maestro som hadde tilsyn med et komplekst orkester. DNA -polymerasene til Luca, langt fra rudimentære, ville ha dyktig dekodede genetiske tegninger, og skapt sammenhengende genetiske fortellinger. Debatten om Luca's sin cellulære struktur vedvarer. Mens noen hevder dens likhet med moderne prokaryote enheter, argumenterer andre for en mobilarkitektur som kan ha vært mer rudimentær, men likevel unikt tilpasset dets primordiale miljø.
Sammenlignende genomikk gir innsikt i den potensielle genomiske rammen for Luca. Denne stamfaderen ville ha huset gener som er sentralt for RNA -syntese, DNA -replikasjon, proteinproduksjon og rudimentære metabolske prosesser. Slike molekylære mekanismer danner grunnlaget for den livlige trivselen til livet vi observerer i dag. En kjernefunksjon som antas å være til stede i Luca's cellulære maskiner, er dens kapasitet for nukleinsyresyntese og vedlikehold. I motsetning til en forenklet modell, ville det genetiske maskineriet til Luca være kompetent, noe som muliggjør DNA -replikasjon og reparasjon, i likhet med en erfaren maestro som hadde tilsyn med et komplekst orkester. DNA -polymerasene til Luca, langt fra rudimentære, ville ha dyktig dekodede genetiske tegninger, og skapt sammenhengende genetiske fortellinger. Debatten om Luca's sin cellulære struktur vedvarer. Mens noen hevder dens likhet med moderne prokaryote enheter, argumenterer andre for en mobilarkitektur som kan ha vært mer rudimentær, men likevel unikt tilpasset dets primordiale miljø.
Kjernen i Luca's eksistens hviler ikke bare i sitt DNA eller RNA, men i dens overordnede biokjemiske maskineri. Se for deg et cellulært system som allerede er utstyrt for å utnytte energi, behandle næringsstoffer og opprettholde genetisk troskap, og forberede grunnlaget for livets påfølgende diversifisering. Utover sin genetiske dyktighet, ville Luca ha navigert over utfordringer som å skaffe energi og opprettholde et internt miljø. De potensielle metabolske traséene den brukte, håpes å tilby vinduer til tidlige tilpasninger. Når det vitenskapelige samfunnet fordyper seg dypere i Luca'ss attributter og betydning, står det som et fyrtårn, og lyser opp livets tidlige grunnlov. å forstå LUCA er å sette pris på et sentralt tidspunkt og utgangspunkt for en evolusjonær fortelling. Mens han ikke representerer livets opprinnelse, avgrenser Luca et rike der grunnleggende molekylære systemer var i drift, banet den vei for den antatte måten for den evolusjonære dansen, som ville ha dannet den mangfoldige biosfæren vi nå observerer.
Bilde 1. Illustrasjon av RNA-reparasjon (O. Grasso)
Primordial fase: gransking av jordens tidlige miljø
Jakten på å forstå livets opprinnelse har pekt det vitenskapelige samfunnet mot forskjellige miljøer som spenner fra dyphavs hydrotermiske ventilasjonssjakter til overflatebasseng rike på prebiotiske forbindelser. Plassert i havets dyp, hydrotermiske ventilasjonssjakter, med sine mineralrike utslipp og temperaturgradienter, blir de postulert som potensielle livskrybber. Disse miljøene tilbyr en blanding av kjemikalier og den nødvendige energien som kan favorisere prebiotiske reaksjoner. Det gjenstår imidlertid flere spørsmål. Hvordan vil delikate organiske molekyler, avgjørende for livet, motstå de intense forholdene og ekstreme trykk på disse dypene? I tillegg kan den raske fortynningen av eventuelle syntetiserte forbindelser i enorme havvolum utfordre akkumulering av kritiske biomolekyler. Konseptet med den primordiale "suppen" fremkaller visjoner om overflatebassenger som vrimler av en blanding av organiske forbindelser, muligens drevet av lyn og ultrafiolett (UV) stråling.
Likevel utgjør den konstante eksponeringen for UV -stråler også et mysterium. Mens de kan katalysere noen reaksjoner, kan de også forringe sensitive organiske forbindelser. Balansen mellom skapelse og ødeleggelse i slike scenarier er gjenstand for granskning. Eksterne hendelser og forhold har vært involvert i muligens å styre traséene som fører til liv. Likevel, var disse hendelsene virkelig deterministiske, og pushet en ikke-levende verden mot livets rand? Meteoritter, lastet med organiske forbindelser, er blitt foreslått som eksterne givere som kan ha frø jord med prebiotiske molekyler. Reisen gjennom jordens atmosfære utsetter imidlertid disse forbindelsene for ekstreme temperaturer. Ville disse meteoritt-forbindelsene virkelig overleve og forbli uendret etter en slik brennende avstamning? Den tidlige jordens atmosfære, som antas å redusere naturen, kan ha lettet syntesen av organiske forbindelser. Men den nøyaktige sammensetningen og dens implikasjoner for dannelse av biomolekyler forblir gjenstander for debatt. Var atmosfæren virkelig gunstig, eller kjemper vi ennå med å forstå nyansene?
Gitt de mange forholdene og hypotesene som er presentert, oppstår spørsmålet: var disse omstendighetene tilstrekkelig deterministiske til å konkludere livets uunngåelige fremvekst? Selv om det er erkjent at visse forhold kan favorisere syntesen av organiske forbindelser, gir det en større kløft fra enkle molekyler til komplekse, funksjonelle cellulære maskiner. Bare en samling av molekyler tilsvarer ikke livet. Hvordan skjedde da spranget fra kjemi til cellulær kompleksitet? Livets maskiner henger sammen med synkroniserte interaksjoner mellom biomolekyler. Orkestrering av disse interaksjonene i fravær av en ledende cellulær ramme, er en betydelig utfordring for vår forståelse. Ved å undersøke de potensielle fødselsplassene i livet og de ytre faktorene som påvirker det, er det tydelig at fortellingen er langt fra fullstendig. Selv om visse forhold kan favorisere syntesen av biomolekyler, forblir progresjonen til et funksjonelt, selvreplikerende system gåtefull. Når vi våger oss videre inn i dette domenet, er det sentralt å nærme oss disse hypotesene med en kritisk linse, sette pris på nyanser og søke klarhet midt i kompleksitetene.
Fra molekyler til liv: Deling inn i gåte med biopolymeroppkomst
 På det mest grunnleggende nivået er livets maskineri drevet av en serie molekyler som fungerer som byggesteiner: aminosyrer, nukleotider og lipider, for å nevne noen. Mens laboratorieeksperimenter har vist at visse basiske molekyler kan dannes under simulerte tidlige jordforhold, er det fortsatt dvelende spørsmål om utbyttet, renhet og stabilitet av disse molekylene. For eksempel produserte Miller-Urey-eksperimentet aminosyrer. Likevel er det verdt å spørre: ble det brukt forhold i slike eksperimenter, som reflekterte den tidlig jorden? Videre, i en prebiotisk setting, uten enzymatisk veiledning, hvordan ble de rette molekylene selektivt syntetisert og opprettholdt midt i en mengde uønskede biprodukter?
På det mest grunnleggende nivået er livets maskineri drevet av en serie molekyler som fungerer som byggesteiner: aminosyrer, nukleotider og lipider, for å nevne noen. Mens laboratorieeksperimenter har vist at visse basiske molekyler kan dannes under simulerte tidlige jordforhold, er det fortsatt dvelende spørsmål om utbyttet, renhet og stabilitet av disse molekylene. For eksempel produserte Miller-Urey-eksperimentet aminosyrer. Likevel er det verdt å spørre: ble det brukt forhold i slike eksperimenter, som reflekterte den tidlig jorden? Videre, i en prebiotisk setting, uten enzymatisk veiledning, hvordan ble de rette molekylene selektivt syntetisert og opprettholdt midt i en mengde uønskede biprodukter?
Naturlig utvalg fungerte ikke i løpet av den prebiotiske tiden på jorden
I det primære miljøet var styrte seleksjonsmekanismer fraværende. Komplekse systemer i moderne organismer produserer byggesteinene i livet. Før livet som vi kjenner det blomstret, var den tidlige jorden et blankt lerret, blottet for den ledende hånden av naturlig utvalg. Se for deg en verden der omhyggelig orkestrerte molekylære symfonier i dag ikke eksisterte. I stedet for presise, bevisste formasjoner av nukleinsyrer og de tjue aminosyrene, eksisterte det et kakofont kaos av molekyler. Innenfor det sofistikerte cellulære maskineriet i det moderne liv, er feilsteg som høyrehendte aminosyrer omhyggelig korrigert. Nukleobaser, fosfolipider og karbohydrater, nå med slike bestemte roller, gikk tapt midt i et hav av utallige molekylære utfordrere. Man må lure på: i en verden som vrimler av kjemisk tilfeldighet, hvordan slo ikke-styrte prosesser gjennom, og plukket ut den molekylære eliten som utgjør livet i dag?
Bilde 2. Samling i større enheter; fra Otangelo Grasso.
Opprinnelsen og organiseringen av livets grunnleggende molekyler
De fire hovedmolekylene i livet - DNA, RNA, proteiner og lipider - tjener som hjørnestein for cellulær funksjon og replikasjon. å forstå deres opprinnelse er avgjørende i vår søken etter å forstå oppstarten av livet på jorden. Det kjemiske miljøet på tidlig jord, uten rammer av cellulært liv og arvelige systemer, manglet en merkbar mekanisme for å velge eller gi preferanse til ett molekyl fremfor et annet. Dette reiser faktisk intrikate spørsmål om hvordan slike sofistikerte og informasjonsbelagte molekyler dukket opp fra et prebiotisk miljø. Begrepet selvorganisering eller autokatalyse, der molekyler spontant stiller seg inn i strukturerte arrangementer eller katalyserer sin dannelse, har blitt postert av mange i det vitenskapelige samfunnet. En slik forestilling vil innebære at under spesifikke forhold har visse molekyler en iboende tilbøyelighet til å organisere eller katalysere reaksjoner som vil føre til livets byggesteiner. Likevel krever fremvekst av molekyler, så informasjonsrike som DNA og RNA, en kontekst der spesifisitet og rekkefølge er avgjørende. Oppstarten av proteiner, med deres enorme utvalg av potensielle aminosyresekvenser og konfigurasjoner, trenger mer enn bare tilfeldig sjanse. Dessuten er lipidmolekylene som danner cellulære barrierer ikke bare enkle hydrofobe kjeder, men er involvert i viktige cellulære funksjoner og signalering. Tatt i betraktning disse kompleksitetene, måtte man utlede at den tidlige jorden måtte presentere forhold der spesifisitet ikke bare var mulig, men sannsynlig. Miljøene i denne primordiale verden, med sin utall av mikro-nisjer, ville måtte tilby et bredt utvalg av fysiske og kjemiske gunstige forhold. Det må tenkes at i et så mangfoldig landskap var dannelsen av livets grunnleggende byggesteiner ikke en isolert hendelse, men en serie hendelser, hver finjustert av de spesielle forholdene i dens nisje. Forestillingen om molekyler med iboende egenskaper som driver dem mot livsfremmende konfigurasjoner antyder et nivå av presisjon og raffinement i designet.
Andrew H. Knoll (2012): Fremveksten av naturlig seleksjon: molekylær seleksjon, prosessen der noen få viktige molekyler tjente nøkkelroller i livets opprinnelse, fortsatte på mange fronter. (Kommentar: Observer den uberettigede påstanden) Noen molekyler var iboende ustabile eller svært reaktive, og derfor forsvant de raskt fra scenen. Andre molekyler ble lett oppløst i havene, og ble derfor fjernet. Fortsatt kan andre molekylære arter ha sekvensiert seg ved å binde seg sterkt til overflater av kjemisk lite hjelpsomme mineraler eller klumpet sammen til tjæremasser med lite bruk i ny biologi. I hvert geokjemisk miljø hadde hver slags organisk molekyl sine pålitelige kilder og sine uunngåelige vansker. I en tid, kanskje i hundrevis av millioner av år, ble en slags molekylær likevekt opprettholdt da den nye tilførselen av hver art ble balansert av tapet. Slik likevekt har visselig vedholdende konkurranse blant molekyler, men systemet utviklet seg ikke. (1)
Kommentar: Livets stoff, som forstått gjennom dagens vitenskapelige analyse, viser en intrikat orkestrering av molekylær maskiner. De grunnleggende bio-molekylene er ikke enkle samlinger av atomer; De er sofistikerte enheter som opererer i meget koordinerte metabolske stier. Når vi tenker på tidlig-jord miljøet, blir det en formidabel utfordring å se for seg hvordan slike spesifiserte, komplekse molekyler vil være lett tilgjengelige. De ville ha trengt å danne seg naturlig, uten formidling av enzymer, og samles i betydelige mengder på et enkelt sted, klare til å bidra til livets fundament.
Bilde 3. Alternativ mekanisme forr bio-diversitet -Fra Otangelo Grasso
 1. Livets intrikate dans henger sammen med en endelig samling av detaljerte bio-molekyler, forent i en universelt anerkjent stevne. Dette kjernekonsortiet omfatter de viktigste byggesteinene: RNA, DNA, aminosyrer, fosfolipider og karbohydrater. Med bemerkelsesverdig presisjon utnytter celler disse elementene gjennom nøye koordinerte metabolske veier, og tilsynelatende fraværende i jordens tidlige stadier. Tatt i betraktning prinsippet om abiogenese, er det viktig at disse bio-molekylene eksisterte i en naturlig forekommende tilstand, dannet uten enzymformidling. Videre ville de ha krevd organisasjon for å legge grunnlaget for begynnelsen av de første levende cellene. En overflod av disse bio-molekylene, konsentrert ved et entall fokuspunkt, ville ha vært essensielt.
1. Livets intrikate dans henger sammen med en endelig samling av detaljerte bio-molekyler, forent i en universelt anerkjent stevne. Dette kjernekonsortiet omfatter de viktigste byggesteinene: RNA, DNA, aminosyrer, fosfolipider og karbohydrater. Med bemerkelsesverdig presisjon utnytter celler disse elementene gjennom nøye koordinerte metabolske veier, og tilsynelatende fraværende i jordens tidlige stadier. Tatt i betraktning prinsippet om abiogenese, er det viktig at disse bio-molekylene eksisterte i en naturlig forekommende tilstand, dannet uten enzymformidling. Videre ville de ha krevd organisasjon for å legge grunnlaget for begynnelsen av de første levende cellene. En overflod av disse bio-molekylene, konsentrert ved et entall fokuspunkt, ville ha vært essensielt.
2. Oppretting med et endepunkt i tankene nødvendiggjør en retningsintensjon. Denne forutsetningen øker utfordringene når man tar for seg naturalistiske forslag om livets opphav. På den eldgamle jorden ville en enorm molekylær vidde ha utelukket enhver systematisk konkurranse eller utvalg, noe som gjorde isolasjon av livs-essensielle molekyler fra ikke-essensielle kolleger tilsynelatende usannsynlig. For å tilskrive hele livets organiserte struktur, dets bevaring og evolusjonære progresjon, fra ikke-levende enheter utelukkende til seleksjon, ser ut til å være en forenklet påstand. å antyde at utvalg er den eneste orkestratoren for livets store symfoni, kan savne nyanser som er viktige for vår forståelse.
3. Fra våre observasjoner er en serie tilfeldige hendelser uten retning svært usannsynlig å produsere komponenter med forskjellige formål, spesielt når du ser på store integrerte systemer. Analogt samles ikke murstein fra råvarer eller posisjonerer seg til å danne vegger. Det er en ledende hånd bak deres formasjon. Tilsvarende oppstår ikke fosfolipider, viktige for cellemembraner, spontant fra sine grunnleggende komponenter. Deres dannelse og påfølgende organisasjon antyder en overordnet veiledning. Spørsmålet som vanker: Kan det være en arkitekt bak dette underverket?
Alan W. Schwartz (2007): Et problem som er kjent for organiske kjemikere er produksjon av uønskede biprodukter i syntetiske reaksjoner. For pre-biotisk kjemi, der målet ofte er simulering av forhold på den pre-biotiske jorden og modelleringen av en spontan reaksjon, er det ikke overraskende - men likevel frustrerende - at de uønskede produktene kan konsumere det meste av startmaterialet og føre til ingenting mer enn en ufravikelig blanding, eller -avfall .. de mest kjente eksemplene på fenomenet kan raskt oppsummeres: selv om Miller-Urey -reaksjonen gir et imponerende sett med aminosyrer og andre biologisk signifikante forbindelser, går en stor brøkdel av startmaterialet inn i en brun, tjære-lignende rest, som forblir ukarakterisert; dvs. avfall. Mens 15% av karbonet kan spores til spesifikke organiske molekyler, ser resten ut til å være stort sett ufravikelig, selv om vi bare fokuserer på de oppløselige produktene, må vi fortsatt takle en ekstremt kompleks blanding av forbindelser. De karbonholdige kondrittene, som representerer en alternativ kilde til startmateriale for prebiotisk kjemi på jorden, og må ha lagt til enorme mengder organisk materiale til jorden på slutten av det sene tunge bombardementet (LHB), tilbyr ikke en løsning på det refererte problemet. Det organiske materialet som er til stede i karbonholdige meteoritter er en blanding av en slik kompleksitet at mye oppfinnsomhet har gått til utforming av passende ekstraksjonsmetoder, for å isolere de viktigste klassene av oppløselige (eller solubiliserte) komponenter for analyse. Uansett hva den nøyaktige arten av en RNA -forløper som kan ha blitt det første selvforklarende molekylet, hvordan kan den kjemiske homogeniteten som virker nødvendig for å tillate denne typen mekanismer til og med komme til å eksistere? Hvilken mekanisme ville ha blitt valgt for inkorporering av tre, eller ribose, eller en bestemt byggestein, i korte oligomerer som senere kan ha gjennomgått kjemisk selektiv oligomerisering? Praktisk talt alle modell pre-biotiske synteser produserer blandinger. (2)
A.G.Cairns-Smith (1985): Det er en av de mest singulære trekkene i biokjemiske enheter at denne enkle konvensjonen er universell. Hvor kom en slik avtale fra? Du ser at ikke-biologiske prosesser ikke som regel viser noen skjevhet på den ene eller den andre måten, og det har vist seg å være spesielt vanskelig å se noen realistisk måte som noen av bestanddelene i en 'prebiotisk suppe' ville ha hatt overveiende 'venstre- Handt 'eller høyrehendt' molekyler. Det er dermed spesielt vanskelig å se denne funksjonen som å ha blitt pålagt av de første betingelsene. ()3
A.G.Cairns-Smith (1985): Genetisk overtakelse, side 70: Anta at ved en tilfeldighet en spesiell coacervate-dråpe i et primordialt hav tilfeldigvis hadde et sett med katalysatorer, etc. som kan konvertere karbondioksid til D-glukose. Ville dette ha vært et stort skritt fremover mot livet? Sannsynligvis ikke. Før eller siden ville dråpen sunket til bunnen av havet og aldri bli hørt om igjen. Det ville ikke ha hatt noe å si hvor genialt eller livslignende et tidlig system var; Hvis det manglet evnen til å gi videre til avkom hemmeligheten bak suksessen, kan det like gjerne aldri ha eksistert. Så jeg ser ikke livet som dukker opp som en selvfølge fra den generelle utviklingen av kosmos, via kjemisk evolusjon, i en stor gradvis prosess med kompleksifisering. I stedet, etter Muller (1929) og andre, ville jeg ta et genetisk syn og se livets opprinnelse som hengende på et ganske presist teknisk puslespill. Hva ville vært den enkleste måten som arvelig maskineri kunne ha blitt dannet på den primitiv jord?(4)
William Dembski (2002): Problemet er at naturen har for mange alternativer og uten design kunne man ikke sortere gjennom alle disse alternativene. Problemet er at naturlige mekanismer er for uspesifikke til å bestemme et spesielt resultat. Naturlige prosesser kan teoretisk danne et protein, men også kompatible med dannelsen av en mengde av andre molekylære samlinger, hvorav de fleste ikke har noen biologisk betydning. Naturen tillater dem full ordningsfrihet. Likevel er det nettopp at frihet som gjør at naturen ikke er i stand til å redegjøre for spesifiserte utfall av liten sannsynlighet. Naturen, i dette tilfellet, i stedet for å være opptatt av å gjøre bare en ting, er åpen for å gjøre mange ting. Likevel når en av disse tingene er en svært usannsynlig spesifisert hendelse, blir design den mer overbevisende, bedre slutningen. Occams barberhøvel koker også ned til et argument fra uvitenhet: I mangel av bedre informasjon bruker du en heuristikk for å akseptere den ene hypotesen over den andre. (5)
Fry, Iris. (2010): Hvor vellykket er RNA-først, RNA-senere, metabolisme-først og forberedende metabolisme-teorier, for å belyse fremveksten av livet som en evolusjonsprosess? Så langt kan ingen av disse paradigmene kreve avgjørende eksperimentell støtte. (6)
Viktige biomolekyler: Utover utvelgelsesområdet
Når det gjelder seleksjonen og spesifisiteten til nukleinsyrer i prebiotisk sammenheng:
1. Renhet av startmolekyler: Midt i en enorm primordial sammenblanding, hvordan dukket den utpregede, kritiske renheten som er nødvendig for RNA og DNA -syntese opp? Slik renhet, med tanke på enormheten og tilfeldigheten til molekylære blandinger som er til stede, peker mot en potensiell veiledningsmekanisme, noe som antyder en overordnet blåkopi kan ha veiledet denne syntesen mot alle odds.
 2. Nucleotid-base seleksjon: I et miljø som vrimler av molekylære kandidater, er det forvirrende hvordan bare fem distinkte nukleotid-basene ble fremtredende. Denne bemerkelsesverdige utvalgspresisjonen, midt i et sannsynlig hav av molekylære kandidater, resonerer med ideen om en prosess som kan ha blitt intelligent koreografert i stedet for et produkt av ren tilfeldighet. Hydrogenbindingsstyrken i DNA er fint innstilt for å sikre stabilitet og spesifisitet i dobbelt helix-formasjonen med adenin sammenkobling med tymin, og guanin sammenkobling med cytosin. Riktig tautomere former for nukleotidbaser er avgjørende for å opprettholde passende hydrogenbindingsmønstre, noe som sikrer nøyaktig replikering og overføring av genetisk informasjon. Mens det eksisterer mange mulige analoge strukturer for nukleotidbaser, er det bare spesifikke baser som brukes i biologiske systemer på grunn av deres unike kjemiske egenskaper og baseparrings-spesifisitet.
2. Nucleotid-base seleksjon: I et miljø som vrimler av molekylære kandidater, er det forvirrende hvordan bare fem distinkte nukleotid-basene ble fremtredende. Denne bemerkelsesverdige utvalgspresisjonen, midt i et sannsynlig hav av molekylære kandidater, resonerer med ideen om en prosess som kan ha blitt intelligent koreografert i stedet for et produkt av ren tilfeldighet. Hydrogenbindingsstyrken i DNA er fint innstilt for å sikre stabilitet og spesifisitet i dobbelt helix-formasjonen med adenin sammenkobling med tymin, og guanin sammenkobling med cytosin. Riktig tautomere former for nukleotidbaser er avgjørende for å opprettholde passende hydrogenbindingsmønstre, noe som sikrer nøyaktig replikering og overføring av genetisk informasjon. Mens det eksisterer mange mulige analoge strukturer for nukleotidbaser, er det bare spesifikke baser som brukes i biologiske systemer på grunn av deres unike kjemiske egenskaper og baseparrings-spesifisitet.
Bilde 4. Adenin kobles med Tymin, og Guanin med Cytosin
3. Strukturell spesifisitet av puriner: Den intrikate to-ring strukturen til puriner, som dukket opp som dominerende er fascinerende, gitt de utallige molekylære konfigurasjonene som er mulig. Puriner, med sin spesifikke dobbeltringstruktur, ble valgt for sin unike evne til å danne stabile og spesifikke hydrogenbindinger med deres komplementære pyrimidiner i DNA, noe som sikrer nøyaktig genetisk informasjonslagring og overføring. Deres spesielle atomarrangement og funksjonelle grupper gir de nødvendige kjemiske egenskapene og baseparringsspesifisiteten avgjørende for integriteten og funksjonen til DNA i biologiske systemer. Denne slående stru kturelle konsistensen, spesielt under primitive forhold, ser ut til å gå inn for en målrettet strukturell blåkopi.
4. Strukturell spesifisitet av pyrimidiner: Hvordan dukket pyrimidiner frem, med sin enkeltring sammensatt av 4 karbon- og 2 nitrogenatomer, som den foretrukne strukturen, gitt de utallige potensielle konfigurasjonene? Pyrimidiner, med sin distinkte enkeltringstruktur, ble valgt fordi de kan etablere spesifikke og stabile hydrogenbindinger med sine komplementære puriner i DNA. Denne spesifikke strukturen og de tilhørende funksjonelle gruppene sikrer presis baseparring, noe som er essensielt for nøyaktig lagring og overføring av genetisk informasjon i levende organismer. Selve fremveksten av slike distinkte strukturelle spesifisiteter antyder en mulig forsettlig molekylær koreografi.
5 Baseseparasjon i komplekse blandinger: Midt i mengden av lignende molekyler som kunne ha dannet seg, hvordan ble de funksjonelle nukleotid-basene til DNA og RNA selektivt isolert? Evnen til å filtrere ut det essensielle fra det ikke-essensielle innebærer et system som favoriserer presisjon.
6 Ribose Ring seleksjon: Hvorfor bruker livet hovedsakelig ribose med sin 5-karbon sukkerring i ryggraden til RNA og DNA, selv om andre konfigurasjoner som 4 eller 6 karbonringer kunne ha dannet seg, men er ikke-funksjonelle? Denne spesifisiteten antyder et optimalisert design.
7 Riktig atomkobling: Hvordan ble det riktige nitrogenatomet i basen og det passende karbonatomet i sukkeret merket og koblet, spesielt når forskjellige tilkoblinger er mulig? Slik nøyaktighet i molekylære bindinger demonstrerer den intrikate utformingen i grunnleggende biokjemi.
8 Homokiralitet i RNA og DNA: I en verden som kan produsere både venstre og høyrehendte molekyler, hvordan kom livet til utelukkende bruke de høyrehendte konfigurasjonene av RNA og DNA, spesielt ribose, i sin D-form? Det konsistente utvalget av visse kirale former over andre, er fortsatt et dyptgående mysterium, noe som antyder en presis orkestrering i dannelsen av livet.
9 Fravær av pre-biotisk seleksjonsmekanisme: Uten livets maskiner på plass, hvordan ville det prebiotiske miljøet ha favorisert dannelsen av RNA- og DNA -molekyler fremfor andre? Mangelen på en naturlig seleksjonsmekanisme før livet presenterer et mysterium når det gjelder å gjøre rede for deres fremvekst.
10 RNAs doble rolle i informasjon og funksjon: Gitt at RNA kan tjene som både en informasjonsbærer (lik DNA) og en katalysator (som proteiner), hvordan kom det til å skaffe seg denne doble rollen, uten en eksisterende seleksjonsmekanisme? Multi-funksjonaliteten til RNA ser ut til å trosse en enkelt, ikke-styrt opprinnelse.
11 Spesifisitet av nukleotid-parring: Adenin går alltid i par med tymin (eller uracil i RNA), og cytosin med guanin. Hvordan utviklet denne svært spesifikke base-parringen seg, uten en mekanisme for å favorisere riktige sammenkoblinger fremfor uriktige?
12 Opprinnelse av replikasjonsmekanisme: Hvordan ble mekanismen for replikasjon av RNA- og DNA -molekyler til, gitt at replikasjonsprosessen i seg selv ser ut til å kreve en form for seleksjon? Uten en feilkorreksjons-mekanisme ville replikasjonens troskap bli kompromittert.
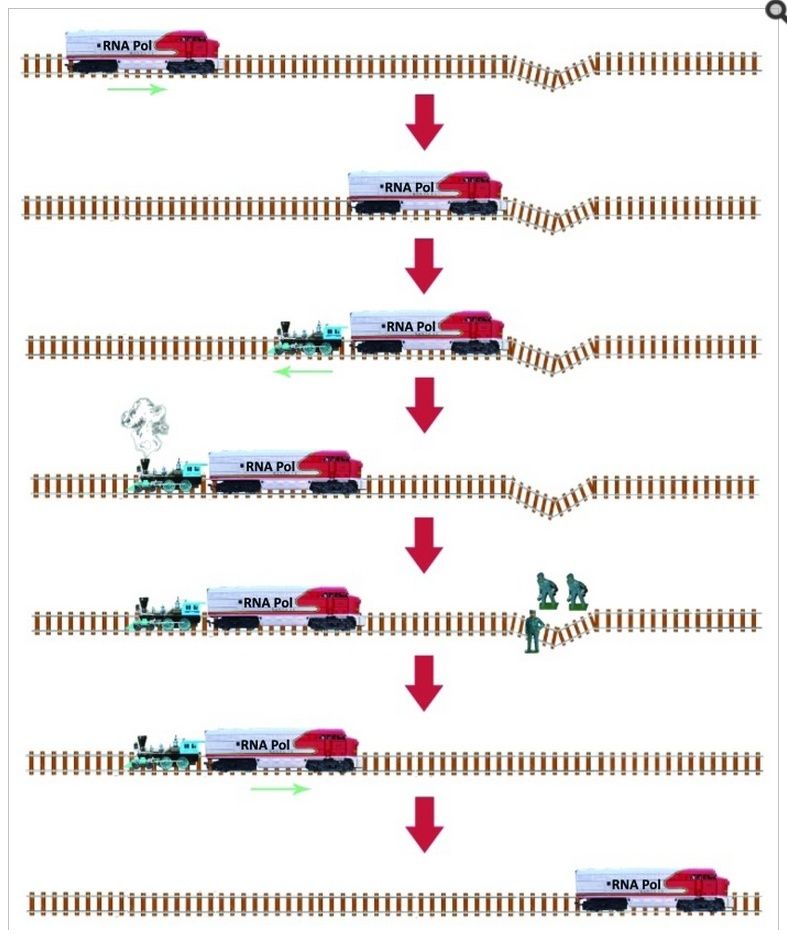
 13 Overgang fra RNA til DNA -verden: Hvis livet begynte med en RNA -verden, som noen antyder, hvordan skjedde overgangen til det mer stabile DNA, spesielt i fravær av pre-biotiske seleksjonsmekanismer for å veilede en slik overgang?
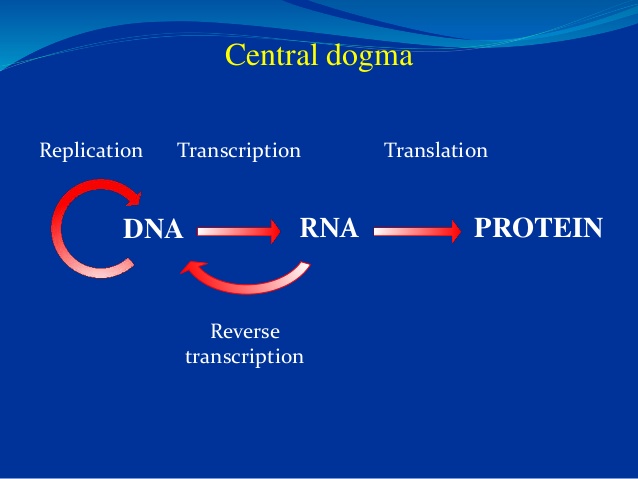
13 Overgang fra RNA til DNA -verden: Hvis livet begynte med en RNA -verden, som noen antyder, hvordan skjedde overgangen til det mer stabile DNA, spesielt i fravær av pre-biotiske seleksjonsmekanismer for å veilede en slik overgang?
Bilde 5. Nåværende sentraldogme: fra DNA via RNA til proteiner
14 Mangel på stabilt pre-biotisk miljø: Gitt den potensielle ustabiliteten og reaktive naturen til tidlig jord, hvordan kan delikate molekyler som RNA og DNA formes og bli bevart uten et stabilt miljø og uten spesifikke mekanismer for å beskytte og favorisere dem?
15 Problem med innledende funksjonalitet: Hvordan fikk det første RNA- og DNA -molekylene den funksjonaliteten som er nødvendig for at de skulle være fordelaktig og underlagt noen form for seleksjon, gitt at de ville trenge å ha et visst nivå av nytteverdi fra begynnelsen?
Utfordringene som er oppregnet, spesielt når de vurderes ut fra deres kumulative effekt, gjør det klart hvorfor livets opprinnelse er et av de mest skremmende og omdiskuterte spørsmålene i både biologi og filosofi. Når vi dykker dypt ned i detaljer av molekylære konfigurasjoner og spesifisiteten som kreves for funksjonelle nukleinsyrer, blir det tydelig at et mangfold av svært spesifikke forhold og hendelser av begivenheter, ville vært nødvendig for at livet dukker opp naturlig. Mange av problemene fremhever den rene spesifisiteten som kreves i flere trinn - fra å velge riktige nukleotidbaser til å danne de riktige bindingene og oppnå funksjonelle sekvenser. Denne spesifisiteten ser ut til å være i strid med tilfeldige, ikke-styrte prosesser.
Det tidlige-jord miljøet var ikke så gunstig for livet som det er i dag. Høy UV -stråling, mangel på beskyttende atmosfære og en mengde reaktive molekyler ville gjøre overlevelse og dannelse av delikate RNA/DNA -kjeder utrolig utfordrende. Betydningen av katalysatorer i moderne biologiske systemer er tydelig. I deres fravær i den pre-biotiske tiden, blir det å oppnå riktig reaksjonshastigheter et annet monumentalt hinder. På dette prebiotiske stadiet var mekanismene for naturlig seleksjon, som leder utviklingen av livsformer, ikke operative. Naturlig seleksjon virker på funksjonelle, replikerende enheter. Hvis du ennå ikke har en funksjonell, replikerende enhet (som de første RNA/DNA -molekylene), kan du ikke ha naturlig seleksjon. Gitt den samlede vekten av disse utfordringene, gjør det ideen om at livet kommer fra rent naturalistiske og upålitelige prosesser svært usannsynlige.
Angående seleksjon og spesifisitet av aminosyrer og proteiner i pre-biotisk sammenheng:
1. Optimalt aminosyrevalg: Gitt at det er over 500 naturlig forekommende aminosyrer på jorden, hvorfor brukes bare 20 av dem spesielt i livets proteiner? Dette unike utvalget ser ut til å gå utover bare sjanse, og peker mot en bevisst ordning, skreddersydd til disse 20 aminosyrenes eksepsjonelle egenskaper.
2. Bi-funksjonell monomerpreferanse: Hvordan har det seg at bifunksjonelle monomerer, de med to funksjonelle grupper, ble dominerende i proteindannelse, når un-ifunksjonelle monomerer kunne forstyrret prosessen? Denne preferansen fremhever viktigheten av struktur i opprettelsen av funksjonelle polymerer.
3. Homo-kiralitets mysteriet: Hvorfor bruker livsformer hovedsakelig venstrehendte aminosyrer og høyrehendte sukkerarter, spesielt når ikke-kirale forløpere kan føre til begge former? Den eksklusive bruken av visse kirale former, taler for et spesifisitet i biologisk design.
 4. Naturlig aminosyrevalg: Med tanke på det store antallet naturlig forekommende aminosyrer, hvordan har det seg da, at livet på jorda først og fremst bruker en konsistent undergruppe for proteinkonstruksjon? Denne konsistensen understreker ideen om et fint innstilt utvalg for optimal funksjon.
4. Naturlig aminosyrevalg: Med tanke på det store antallet naturlig forekommende aminosyrer, hvordan har det seg da, at livet på jorda først og fremst bruker en konsistent undergruppe for proteinkonstruksjon? Denne konsistensen understreker ideen om et fint innstilt utvalg for optimal funksjon.
5. Unngåelse av uønskede biprodukter: Hvilken mekanisme sikrer at syntesen av biologiske aminosyrer ikke resulterer i uønskede eller irrelevante biprodukter? Presisjonen i denne prosessen antyder et høyere orkestreringsnivå.
6. β, γ, δ aminosyre filtrering: Hvordan ble ikke-alfa aminosyrer filtrert ut i den primordiale suppen, gitt deres potensielle tilstedeværelse? Denne filtreringen antyder en prosess som prioriterer funksjonalitet og effektivitet.
Bilde 6. Videre i livets gang: Fra peptider til proteiner, Fra Otangelo Grasso.
7. Adressering av homo-kiralitet: Hvordan kan et miljø med ikke-kirale forløpere, utelukkende føre til venstrehendte aminosyrer? Dette fenomenet er fortsatt ett av de store mysteriene i biokjemi.
8. NH3 -akkumulering: Hvordan ble ammoniakk, essensielt for aminosyresyntese, samlet på den tidlige jorden, til tross for den korte levetiden på grunn av fotokjemisk dissosiasjon? Dette antyder at forholdene på en eller annen måte var optimale for fremvekst av livet.
9. Organosulfur sammenesetnings-dannelse: Hvordan genererte tidlig-jord prosesser organosulfur-forbindelsene som er nødvendige for visse aminosyrer, med tanke på svovelstilstanden i naturen? Eksistensen av disse forbindelsene innebærer en vei som bidrar til livet.
10. Enzymatisk overgang: Hvordan skjedde overgangen fra prebiotisk enantiomer -seleksjon til enzymatiske reaksjoner som transaminering, gitt dens nødvendighet for livets fremvekst? Denne intrikate overgangen taler for kompleksiteten og finessen i biologiske systemer.
11. Aminosyrenes egnethet: Hvordan hard et seg at de valgte aminosyrene er perfekt egnet for å danne strukturer med trange kjerner, noe som muliggjør bindende lommer i proteiner? Den optimale naturen til disse aminosyrene antyder målrettet design.
12. Unikt aminosyresett: Hvorfor ser settet med aminosyrer, som brukes av livet, ut til å være nesten optimal i egenskaper som størrelse, ladning og hydrofobisitet, spesielt sammenlignet med millioner av alternativer? Denne optimaliseringen antyder et design rettet mot maksimal funksjonalitet.
 13. Fremveksten av synteseforordning: Hvordan dukket reguleringen av aminosyre-synteseveier opp, og sikret at tilbudet oppfyller etterspørselen? Slik regulering indikerer et system som ikke bare er sammensatt, men også tilpasningsdyktig.
13. Fremveksten av synteseforordning: Hvordan dukket reguleringen av aminosyre-synteseveier opp, og sikret at tilbudet oppfyller etterspørselen? Slik regulering indikerer et system som ikke bare er sammensatt, men også tilpasningsdyktig.
14. Metabolsk veiovergang: Hvordan skjedde overgangen fra prebiotisk syntese til metabolsk-sti-basert aminosyresyntese, spesielt gitt de mange enzymer som er involvert? Denne komplekse overgangen understreker vanskeligheter i utviklingen av livets systemer.
Bilde 7. Konklusjon om livets opprinnelse
Fra valg av riktige byggesteiner (aminosyrer) til dannelse av komplekse tredimensjonale strukturer (proteiner), gir hvert trinn betydelige utfordringer for naturalistiske forklaringer på livets opprinnelse. Disse problemene krever ikke bare svar for individuelle trinn, men nødvendiggjør også en forståelse av hvordan disse trinnene kan komme sammen, i rekkefølge og synergistisk. Mens det pågår forskning innen abiogenese og noen skritt er gjort, fører den kumulative vekten av disse utfordringene ofte til intense debatter blant forskere, filosofer og teologer angående livets opprinnelse.
Oversatt via google-oversetter - og bilder, ved Asbjørn E. Lund