 Selv-organisering i en etter-Darwinistisk verden
Selv-organisering i en etter-Darwinistisk verden
(Fra Darwins Doubt; S.C.Meyer, Harper One, 2013, Ch.15)
I 2009 publiserte Simon Conway Morris en artikkel, der han hevdet flg.: "når det kommer til den tilsynelatende plutselige tilsynekomsten av dyre-fossiler (i Kambrium), så har uløste problemer åpnet veien til en etter-Darwinistisk verden." (1) Bevis for en virkelig eksplosjon av dyre-fossiler vs. en tilsynelatende, var bakgrunnen for hvorfor biologer har begynt å gå inn i en slik verden.
Året før møttes 16 innflytelsesrike evolusjons-biologer i Altenberg, Australia for å utforske den framtidige kursen til evolusjonsbiologien. Disse biologene hadde mange ulike , og delvis motstridende ideer om hvordan nye livsformer hadde opprunnet. Paleontolgene Douglas Erwin og James Valentine konkluderte også med at eksplosjonen av dyrefossiler i Kambrium var og ble mystisk. Faktisk har evolusjons-biologer i over to tiår allerede, jobbet med hvordan evolusjonære mekanismer med mer kreativ kraft enn mutasjoner og naturlig utvalg, kombinert med arvelighet av disse, kunne formuleres. Felles for dem har vært at de forsøker besvare det stadig viktigere spørsmålet: 'Etter neo-Darwinismen, hva da?'
Som kjent bygger den neo-darwinistiske mekanismen på tre kjerne områder: i) Evolusjons forandring skyldes resultatet av tilfeldige mutasjoner. ii) Naturlig seleksjon siler blant disse forandringene og mutasjonene, slik at noen organismer favoriseres framfor andre. iii) Favoriserte variasjoner må vedvarende arves i påfølgende generasjoner organismer. Virkningen av det påvirker populasjonen til å endres over tid. Disse kalles av noen for de tre pilarene for neo-darwinistisk evolusjon.
Blant de evolusjons-biologene som nå betviler ortodoks neo-darwinistisk teori, stiller de seg enten spørrende eller avvisende til én eller flere av disse tre pilarene. Eldredge and Gould betvilte 'gradualismen', som gjorde at de benektet ideen om at mutasjonsendringer opptrer i små, gradvise endringer. En ny klasse evolusjonære teorier demper betydningen av tilfeldige mutasjoner og vektlegger i stedet noe de kaller 'selvorganisering'.

Bilde 1. Trilobit fossiler
Disse advokatene for selvorganisering foreslår at biologiske former ofte plutselig oppstår av seg selv, ute i naturen. Dette skal skje som en konsekvens av naturlige 'form-lover'. Så vil videre naturlig seleksjon virke for å bevare disse spontant oppståtte ordener. De mener slike spontane, selv-organiserende former gir den ultimate kilden for nye biologiske former. Slik toner de ned betydningen til mutasjoner, og mutasjoner som 'foredles videre' ved naturlig seleksjon.
I 1993 utga den mest framtredende forskeren ved Santa Fe instituttet, biokjemiker Stuart Kaufmann en etterlengtet avhandling: 'The Origin of Order: Self-Organisation and Selection in Evolution' 'Opprinnelsen til Orden: Selvorganisering og seleksjon i evolusjon'. Han ga uttrykk for en skarp kritikk av de skapende kreftene i mutasjons- og seleksjons- mekanismene. Kaufmann peker på viktigheten til strukturell og 'posisjonell' informasjon i celler og celle membraner, som en viktig faktor til å bestemme hvordan ulike celler organiseres inn i ulike dyrs kroppsform. Ved å anerkjenne viktigheten til slike epigenetisk informasjon til dyrs utvikling, avviser også Kaufmann den neo-darwinistiske formodningen, at et genetisk program helt og holdent bestemmer dyrs utvikling.
Kaufmann synes stilltiende å innrømme vansken ved å fremkalle biologisk spesifisitet, bare ut fra kjemiske reaksjoner. I kritikk av sin egen modell, meddeler han at mønstre av molekylær diffusjon, framkalt ved kjemisk katalyse, avgjørende ville avhenge av 'initialbetingelsene'. Med andre ord, det å få et biologisk informasjonsrikt arrangement av kroppsbyggende proteiner, krever at en starter med meget spesifikke (fortrinnsvis informasjonsrike) arrangement av 'autokatalyserende' molekyler.
Hvor denne informasjonen kom fra, sier ikke Kauffman noe om, men det er det avgjørende spørsmålet i livets historie. Kauffman innrømmer at programmereren fininnstiller systemet for å holde det unna de to grøftene: a) å danne en ekstremt rigid orden eller b) å utvikle seg mot kaos. (Sml. 2. termodynamiske lov)
Denne fininnstillingen måtte antageligvis innbefatte en intelligent programmerer, som valgte visse parameterverdier og forkastet andre. Dette innebærer input av informasjon. Cellens orden synes å oppstå fra dynamikken ved genomets nettverk, d.v.s. fra preekistente, uforklarlige kilder ved genetisk informasjon. Kauffmans simuleringer gjør 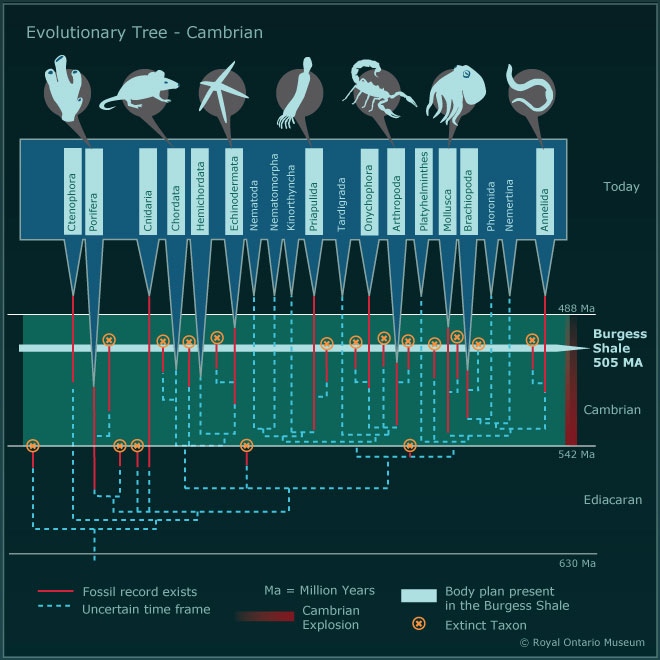 dessuten ikke rede for framvekst av kompleks, spesifikk informasjon.
dessuten ikke rede for framvekst av kompleks, spesifikk informasjon.
Bilde 2. Den plutselige økning i antall arter
Vi viser her et eksempel på hva kompleks, spesifikk informasjon innebærer:
i) Orden: Na-Cl-NA-Cl-Na-Cl-NA-Cl-Na-Cl-NA-Cl-Na-Cl-NA-Cl
ii) Kompleksitet: AZFRT-MPGRTSJHKLKYR
iii) Spesifisert kompleksitet: 'Tid og tidevann venter ikke på noe menneske'.
Den lovliknende, selvorganiserings-prosessen som genererer den slags orden som tilfelle i) representerer i et krystall eller virvelstrøm; generer ikke komplekse sekvenser eller strukturer. Ennå mindre danner de spesifisert kompleksitet, den slags systematikk som er tilstede i et gen eller funksjonelt komplekst organ.
De informasjonsrike sekvensene i DNA, RNA og proteiner karakteriseres verken ved enkel orden (i) eller kun ved kompleksitet (ii). I slike sekvenser er det irregulære og uforutsigbare arrangementet av tegnene (eller bestanddelene) kritiske for den funksjonen sekvensen skal utføre. I slike situasjoner kreves spesifisert kompleksitet: Informasjon med en innebygd kodet betydning, som gir mening for mottager av informasjonen.
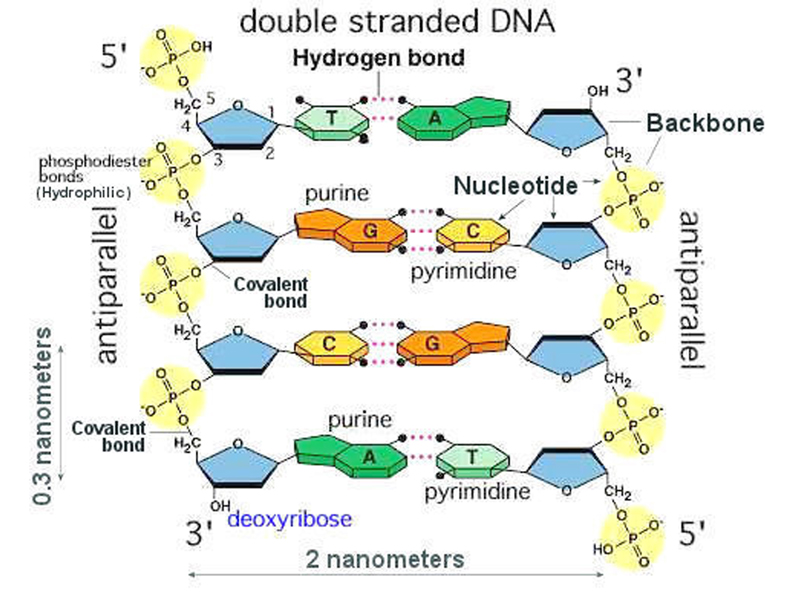
Ellers ville nukleotide-basene repeteres rigid, slik som: ACACACACACACACACACACACAC -noe som ikke ville tillate DNA å lagre eller inneholde spesifisert kompleks informasjon. Et merkelig trekk ved kjemien i DNAet, tillater hvilket som helst av de fire basene (A,C,G og T) å binde til hvilket som helst sted på sukkerfosfatstrengen i DNA-molekylet. Mange folk synes å tro nettopp det at det er kjemiske bindinger i sukkerfosfatstrengen, som utgjør en affinitet som skulle tilsi noen form for selvorganisering. Men, det er ikke påvist at naturalistiske metoder er i stand til å konstruere DNA-språket: Det er ingen kjemiske preferanser knyttet til hvordan noen av nitrogenbasene binder seg til sukkerfosfatstrengen. Et poeng til å ta med seg her, er at sukker ikke kan produseres samtidig med at nitrogen forekommer. Flg. dilemma oppstår da: Enten forekommer aminosyrer i omgivelsene, og da kan ikke sukker produseres. Eller så forekommer ikke aminosyrer, og da blir dannelse av proteiner umulig.
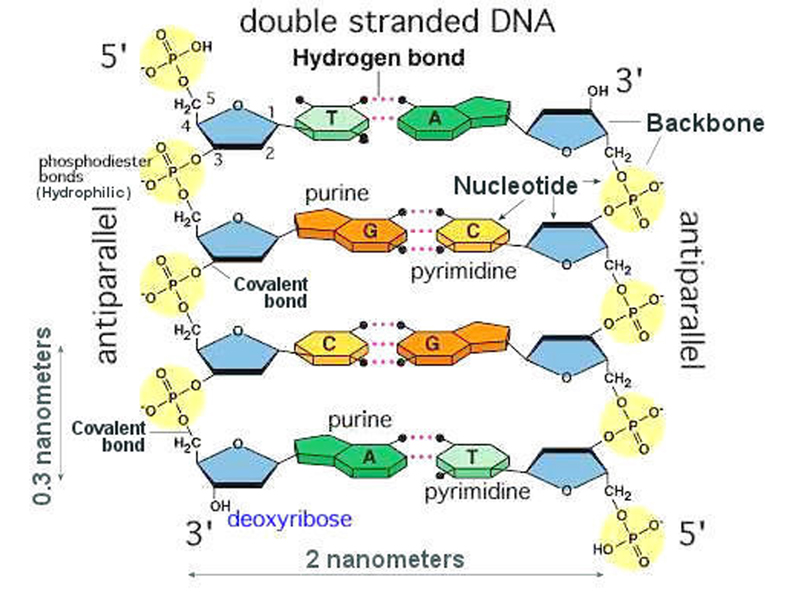 Bilde 3. Sukker-fosfatstreng i DNA
Bilde 3. Sukker-fosfatstreng i DNA
Hubert Yockey, lenge en ledende kapasitet i anvendelse av informasjonsteori på molekylær biologi, var den første til å erkjenne problemet med selvorganisering av biologisk informasjon. I følge ham falt disse teoriene av to grunner: a) de skilte ikke orden fra informasjon og b) informasjonen i DNA-molekylet stammer typisk, ikke fra lov-like prosesser krefter som tiltrekning eller adhesjon. Han uttrykte i 1977: "makromolekyler kan kode genetiske beskjeder, og derfor bære informasjon fordi sekvensen av baser knapt blir påvirket i det hele av (selvorganiserende) fysio-kjemiske krefter." (2)
I det hele er selvorganiserende strukturer ekstremt beskjedne i innhold. De inkluderer repeterende atommønstre i krystaller, enkle geometriske figurer, linjeformer, triangler, virvelstrømmer eller strømninger, av type (i) evt (ii). Ingen av dem innehar den spesifiserte kompleksiteten (iii) som innehas av DNA og RNA. Det gjør heller ikke de komplekse arrangement av proteiner, celler, vev eller organer som er nødvendig for å bygge en fungerende form for dyreliv.
Referanser:
1. Simon Conway Morris: Current Biology; 'Walcott, the Burgess Shale and the rumours of a post-Darwinian World; by . R928
2. Yockey, “A Calculation of the Probability of Spontaneous Biogenesis by Information Theory”, s. 380.
Oversettelse samt utvalg av stoff og bilder ved Asbjørn E. Lund
{kind=link}