Opprinnelsen til kroppsdannelse
Fra 'Darwin's doubt' (Kap. 13)
 I 1995 ble Nobel prisen i medisin gitt til to tyske forskere (Wieshaus og Nüsslein-Vollhard) for deres nye og grundige arbeid med å kartlegge genomet til bananfluer. De hadde i årtier arbeidet med mutasjonseksperimenter på disse, med nedslående resultater. De hadde nok framkalt forandringer i form av ekstremiteter, dupliseringer og endog noe funksjonsendring. Men ingenting hadde fungert i helheten. Riktignok var balanseorganer blitt endret til vinger, bare med den følge at flua mistet styringen.. Problemet var i følge Wieshaus at de trodde de hadde funnet alle genene nødvendig for å endre krop
I 1995 ble Nobel prisen i medisin gitt til to tyske forskere (Wieshaus og Nüsslein-Vollhard) for deres nye og grundige arbeid med å kartlegge genomet til bananfluer. De hadde i årtier arbeidet med mutasjonseksperimenter på disse, med nedslående resultater. De hadde nok framkalt forandringer i form av ekstremiteter, dupliseringer og endog noe funksjonsendring. Men ingenting hadde fungert i helheten. Riktignok var balanseorganer blitt endret til vinger, bare med den følge at flua mistet styringen.. Problemet var i følge Wieshaus at de trodde de hadde funnet alle genene nødvendig for å endre krop psplanen til Drosophila (typen bananflue). Likevel er ikke resultatene lovende som råmateriale for makro-evolusjon. Det neste spørsmålet er hva som ville være de rette mutasjoner for makro-evolusjon. Og vi vet ikke svaret på det..
psplanen til Drosophila (typen bananflue). Likevel er ikke resultatene lovende som råmateriale for makro-evolusjon. Det neste spørsmålet er hva som ville være de rette mutasjoner for makro-evolusjon. Og vi vet ikke svaret på det..
30 år senere vet fortsatt ikke utviklings -og evolusjons-biologien svaret på det spørsmålet. På samme tid har mutasjonsgenetiske forsøk reist brysomme spørsmål angående rollen til mutasjoner i opphavet til utforming av dyrekropper. Det gjelder for forsøk på en rekke små dyrearter. Den neo-darwinistiske mekanismen har feilet i å forklare opphavet til nye gener og proteiner, nødvendig for at nye organer/kroppsdeler kan oppstå. Men selv om de kunne det, gjenstår et mye mer fundamentalt problem: For å bygge et nytt dyr og etablere kroppsplanen for det, trengs at proteiner arrangeres i høyere ordens strukturer. Med en gang nye proteiner oppstår, må 'noe' arrangere dem til å ta sin del i distinkte celle-typer. Disse må igjen organiseres, for å danne distinkte cellevev, organer og kropps-planer. Denne organiserings-prosessen foregår under utviklingen av embryos.
 Utviklingsbiologien kaller prosessen der embryoet utvikler seg til modne organismer (gjelder flercellede organismer) for ontogeni. Feltet har stadig økt vår forståelse for hvordan kroppsplaner oppstår. Mye av kunnskapen har kommet gjennom såkalte modell-systemer, organismer som biologer lett kan mutere i laben (bananfluen Drosophila, samt en rundorm etc.) Selv om det er forvirrende ulikheter, er det ett felles trekk med all ontogeni: Den starter med ei celle og slutter med mange ulike celler. I de fleste arter starter det med ett befruktet egg. Med en gang egget deler seg i dets datter-celler, blir egget et embryo. Da begynner det å stile mot et vel-definert mål, nemlig en voksen form som kan reprodusere. For å nå det målet må embryoet produsere mange spesialiserte celle-typer på korrekt sted og tid.
Utviklingsbiologien kaller prosessen der embryoet utvikler seg til modne organismer (gjelder flercellede organismer) for ontogeni. Feltet har stadig økt vår forståelse for hvordan kroppsplaner oppstår. Mye av kunnskapen har kommet gjennom såkalte modell-systemer, organismer som biologer lett kan mutere i laben (bananfluen Drosophila, samt en rundorm etc.) Selv om det er forvirrende ulikheter, er det ett felles trekk med all ontogeni: Den starter med ei celle og slutter med mange ulike celler. I de fleste arter starter det med ett befruktet egg. Med en gang egget deler seg i dets datter-celler, blir egget et embryo. Da begynner det å stile mot et vel-definert mål, nemlig en voksen form som kan reprodusere. For å nå det målet må embryoet produsere mange spesialiserte celle-typer på korrekt sted og tid.
 Under ontogenien dobles antall celler hele tiden, ofte i en rasende fart. Antall celle-delinger og totalt antall celler reflekterer tallet på ulike celle-typer den modne organismen trenger. Dette innebærer igjen å produsere ulike proteiner for ulike celle-typer. Hvert protein må differensieres, fordi de gjerne utfører helt ulike oppgaver. Under ontogenien må da de rette genene bli slått av/på for å sikre at rett protein blir produsert til rett tid i de rette celle-typer. Spesifikke proteiner spiller aktive roller i regulering hvordan gener framtrer for å bygge andre proteiner. Proteinene som spiller denne rollen er kjent som transkripsjons-regulatorer(TRs) eller transkripsjons-faktorer (TFs). Disse knytter seg direkte til spesifikke steder i DNA og enten hindrer eller aktiver transkripsjon av spesifikke gener inn i RNA. De inneholder instruksjoner om hvilke gener de skal slå av eller på. Den tredimensjonale geometrien utøver karakteristiske trekk ved DNA-binding: inkl. et område av 61 aminosyrer som pakker seg omkring den doble DNA-spiralen.
Under ontogenien dobles antall celler hele tiden, ofte i en rasende fart. Antall celle-delinger og totalt antall celler reflekterer tallet på ulike celle-typer den modne organismen trenger. Dette innebærer igjen å produsere ulike proteiner for ulike celle-typer. Hvert protein må differensieres, fordi de gjerne utfører helt ulike oppgaver. Under ontogenien må da de rette genene bli slått av/på for å sikre at rett protein blir produsert til rett tid i de rette celle-typer. Spesifikke proteiner spiller aktive roller i regulering hvordan gener framtrer for å bygge andre proteiner. Proteinene som spiller denne rollen er kjent som transkripsjons-regulatorer(TRs) eller transkripsjons-faktorer (TFs). Disse knytter seg direkte til spesifikke steder i DNA og enten hindrer eller aktiver transkripsjon av spesifikke gener inn i RNA. De inneholder instruksjoner om hvilke gener de skal slå av eller på. Den tredimensjonale geometrien utøver karakteristiske trekk ved DNA-binding: inkl. et område av 61 aminosyrer som pakker seg omkring den doble DNA-spiralen.
Transkripsjons regulatorer og faktorer (TRs og TFs) blir selv kontrollert av komplekse kretser og signaler, overført av andre gener og proteiner. Alt har en overveldende kompleksitet og presisjon. Mange av de regulerende gener som spiller nøkkel-roller i å omforme embryoer til modne organismer, er avdekket av forskningen til Wieshaus og Nüsslein-Vollhard. Denne forskningen avslørte også en fundamental vanskelighet, som gikk rett til 'hjertet' på det neo-Darwinistiske synet på livet.
 Mutasjoner tidlig i embryo-stadiet
Mutasjoner tidlig i embryo-stadiet
Normal utvikling for alle dyr kan representeres som et ekspanderende beslutnings-nettverk der de tidligste hendelser har større betydning enn senere. Regulerende gener og deres DNA-bindende protein kontrollerer dette utfoldende nettverket. Hvis da ett regulerende protein blir endret eller ødelagt av mutasjoner, så sprer effekten seg nedover i hele nettverket. Vanskeligheten med å forsøke endre en organisme til en annen, ligger i vanskeligheten ved å resette et antall av de mange kontrollerende svitsjene, slik at det tillater individets kroppsutvikling-uten å ødelegge det.
Det bare mutasjoner framkalt tidlig i embryo-stadiet som har noen sjanse til å avstedkomme storskala, makroevolusjonær forandring. Sentvirkende mutasjoner kan dermed ikke framkalle store endringer. Dette bekreftes av flere evolusjonær-biologer (G. Miklos, K.Thompson ) Likevel i eksperimenter fra tidlig på 1900-tallet (T.H.Morgan..), så har erfaringen ensidig vært ødeleggende for organismer utsatt for mutasjon, inntil i dag.. Slike mutasjoner er enten dødelige eller resulterer i organismer som ikke kan leve i vill tilstand (neo-darwinist pioner R.A.Fisher) En endring tidlig i embryo-stadiet, ville kreve en rekke andre koordinerte forandringer i atskilte, men funksjonelt inter-relaterte utviklingsprosesser. Det er den tette integreringen av funksjoner som gjør at tidlige mutasjoner fører til deformering eller død i embryo-stadiet.
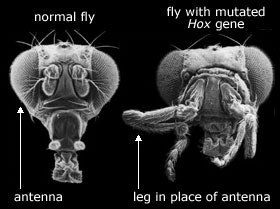 [J. Schwarz argumenterer i boka 'Sudden Origin' for at Hox-gener kan våre ansvarlige for plutselige endringer i fossil-registeret (1). Men det er mange gode grunne til å betvile denne påstanden.
[J. Schwarz argumenterer i boka 'Sudden Origin' for at Hox-gener kan våre ansvarlige for plutselige endringer i fossil-registeret (1). Men det er mange gode grunne til å betvile denne påstanden.
i) Nettopp fordi hox-gener koordinerer uttrykket av så mange andre ulike gener, har eksperimentelt fremkalte mutasjoner i hox-gener vist seg skadelige. Dette bekreftes f.eks. av McGinnis og Kurziora: "de fleste mutasjoner i homeotiske (hox)-ganer forårsaker fatale fødsels-skader." (3) Ved mutasjon av hox-gener hos bananfluer, har biologer fremkalt den dramatiske Antennapedia-mutanten, en ulykkelig flue der bena gror ut fra hodet, der 'antennen' skulle våre. (4) Antennapedia kan ikke overleve i naturen; den har vanskelig for å reprodusere seg og dets avkom dør lett.
ii) Hox-gener i alle dyreformer er uttrykt ETTER at kroppsplanen er etablert. I bananfluer, da hox-gener uttrykkes, er allerede seks tusen celler dannet, og basis geometri for flua, dens fram-bakover akse og rygg-buk akser, er allerede godt etablert. Av den grunn kan ikke og bestemmer ikke hox-gener dannelse av kroppsplan. E. Davidson og E. Davidson (foregangsmenn på området) har påpekt at hox-gener inntreffer mye senere i embryo-perioden enn globale kroppsplan-spesifikajoner, selv om de er nødvendige for korrekt regional eller lokal differensiering innenfor en kroppsplan. Primår opprinnelse til kroppsplan avgjøres av fremdukking av mye dypere kontroll-elementer. Davidson kaller dem kjerne 'utviklings gen-regulerende nettverk' dGRNs (5). å endre slike dGRNs har uten unntak ført til at embryo-utvikling i dyr har stoppet opp.
iii) Hox-gener forsyner bare informasjon for å bygge proteiner som fungerer som svitsjer som slår gener av og på. Det er genene som de regulerer som inneholder informasjon for å bygge proteiner, som former deler til andre strukturer og organer. Hox-genene selv inneholder ikke informasjon til å bygge disse strukturelle delene. M.a.o. hox-gener har ikke all nødvendig informasjon for å bygge nye vev, organer eller kroppsdeler. Således kan ikke det å mutere disse muligens kunne generere nye former for dyreliv.
 Teoretisk biolog E. Szathmary bemerker at Hox-gener ikke koder for proteinene som kroppsdeler er laget av. Det følger som han insisterer at mutasjoner i Hox-gener ikke av seg selv kan bygge nye strukturer.(7) Han betviler også hvorvidt mutasjoner i disse genene har mye kreativ kraft. (29) Talsmenn for evolusjonåre utviklingsbiologer har forlatt ideen at små, inkrementelle variasjoner og mutasjoner er tilstrekkelige for å produsere nødvendige evolusjonåre endringer. I stedet har de gjennopplevd forestillingen om makro-mutasjoner (teorien om 'håpefulle monstre'). De har da også gjenopplivet et gammelt problem: større mutasjonsendringer er ikke levedyktige, og levedyktige mutasjonsendringer er ikke større. Således har ikke evolusjonår utviklingsbiologi lyktes i å gjøre rede for opprinnelsen til ny biologisk informasjon. -Avsnitt om hox-gener er hentet fra 'Theistic Evolution - a Scientific, Philosophical and Theological critique, kap.8]
Teoretisk biolog E. Szathmary bemerker at Hox-gener ikke koder for proteinene som kroppsdeler er laget av. Det følger som han insisterer at mutasjoner i Hox-gener ikke av seg selv kan bygge nye strukturer.(7) Han betviler også hvorvidt mutasjoner i disse genene har mye kreativ kraft. (29) Talsmenn for evolusjonåre utviklingsbiologer har forlatt ideen at små, inkrementelle variasjoner og mutasjoner er tilstrekkelige for å produsere nødvendige evolusjonåre endringer. I stedet har de gjennopplevd forestillingen om makro-mutasjoner (teorien om 'håpefulle monstre'). De har da også gjenopplivet et gammelt problem: større mutasjonsendringer er ikke levedyktige, og levedyktige mutasjonsendringer er ikke større. Således har ikke evolusjonår utviklingsbiologi lyktes i å gjøre rede for opprinnelsen til ny biologisk informasjon. -Avsnitt om hox-gener er hentet fra 'Theistic Evolution - a Scientific, Philosophical and Theological critique, kap.8]
dGRNs (development Gen Regulatory Networks)
En annen linje i utviklingsbiologi har åpenbart en relatert utfordring til den kreative kraften i den neo-Darwinske mekanismen. Utviklings-biologer har oppdaget at proteiner og RNA, som trengs for utvikling av kropps-utformingen til dyr, overfører signaler som influerer måten celler utvikler og differensieres. Disse signal-molekylene influerer hverandre til å danne nettverk av koordinert interaksjon. For eks. avhenger utskilling av signal-molekyl av når et signal mottas fra et annet molekyl. Og så påvirker det overføringen av enda flere, som alle er koordinerte og integrerte, for å utførte spesifikke tids-kritiske funksjoner.
 Eric Davidson er den biologen som mest dyptgående har undersøkt regulerings-logikken i dyrs utvikling. Davidson observerte at cellene til et individuelt dyr, samme hvor variert i form eller funksjon, generelt besto av identiske genomer. Gjennom livssyklusen til en organisme, så uttrykte genomet til disse spesialiserte cellene bare en liten del av sitt DNA til en gitt tid, og produserte dermed ulikt RNA som resultat. Dette faktum antyder sterkt at det finnes ett genetisk kontrollsystem for hele organismen, som skrur brytere på og av etter hvor en er i embryo-utviklingen. Likeså at et slikt system styrer utviklingen fra egg til moden organisme, i det ulike celle typer dannes. Britten og Davidson la fram en teori ang. dette i 1969. Selv om lite da var kjent, deduserte de at et slikt system måtte være virksomt. Det var for det første fordi hundrevis av spesialiserte celler oppsto gjennom utviklingen av dyr, fra embryo til fullt utviklet. For det andre at hver celle inneholdt samme genom, så sluttet de at et slags kontroll-system måtte bestemme hvilke gener som uttrykkes i ulike celler til ulike tider, for å sikre differensiering av ulike celletyper fra hverandre. Derfor måtte det være en regulerende logikk som overstyrte og koordinerte, hvordan genomet uttrykte seg til enhver tid.
Eric Davidson er den biologen som mest dyptgående har undersøkt regulerings-logikken i dyrs utvikling. Davidson observerte at cellene til et individuelt dyr, samme hvor variert i form eller funksjon, generelt besto av identiske genomer. Gjennom livssyklusen til en organisme, så uttrykte genomet til disse spesialiserte cellene bare en liten del av sitt DNA til en gitt tid, og produserte dermed ulikt RNA som resultat. Dette faktum antyder sterkt at det finnes ett genetisk kontrollsystem for hele organismen, som skrur brytere på og av etter hvor en er i embryo-utviklingen. Likeså at et slikt system styrer utviklingen fra egg til moden organisme, i det ulike celle typer dannes. Britten og Davidson la fram en teori ang. dette i 1969. Selv om lite da var kjent, deduserte de at et slikt system måtte være virksomt. Det var for det første fordi hundrevis av spesialiserte celler oppsto gjennom utviklingen av dyr, fra embryo til fullt utviklet. For det andre at hver celle inneholdt samme genom, så sluttet de at et slags kontroll-system måtte bestemme hvilke gener som uttrykkes i ulike celler til ulike tider, for å sikre differensiering av ulike celletyper fra hverandre. Derfor måtte det være en regulerende logikk som overstyrte og koordinerte, hvordan genomet uttrykte seg til enhver tid.
Gjennom de siste to tiår har forskning i genomet avslørt at ikke-kodende regioner av genomet, kontrollerer og regulerer timingen til hvordan protein-kodende regioner av genomet uttrykker seg. Sammen fungerer ikke-protein kodende og protein-kodende regioner av genomet som et nettverk, i følge Davidson. Davidson valgte fra 1971 et sjøpiggsvin (Strongylocentrotus purpuratus) som modell for sitt eksperiment. Sammen med kolleger utviklet Davidson et pioner-prosjekt og eksperimentelle protokoller som krevdes, for å dissekere og kartlegge sjøpiggsvinets genetiske regulerende nettverk.
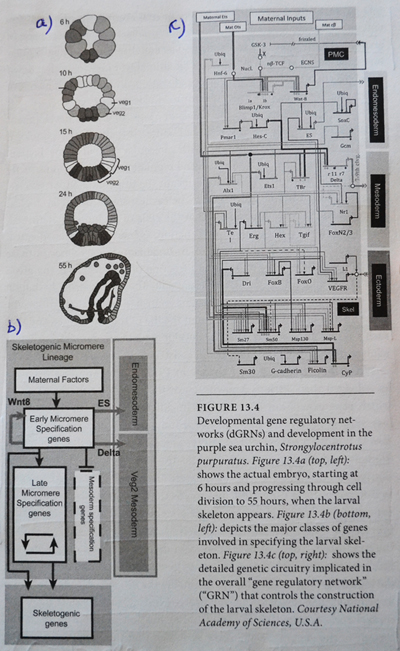
Den fantastiske kompleksiteten de fant framstilles på figure 13.4 her ved siden. Del a viser utvikling av embryoet i en alder fra 6 til 55 timer. Øverst er det 4 runder med celledeling og 16 celler (2 opphøyd i 4). I de neste fire stadiene ser vi en tiltagende spesialisering i kroppsformen. Del b er et skjematisk diagram over hovedklassene av gener og cellevev, i løpet av embryo-utviklingen. Sammenhengen markeres med kontrollpiler. Fig 13.c viser 'det genetiske kretskortet' (i følge Davidson) som skrur på de spesifikke genene, som igjen produserer det nødvendige strukturelle proteinet for å danne sjøpiggsvinet.
For å uttrykke genene som lager proteiner for å danne skjelettet, så må først gener som aktiveres timer tidligere først spille sin rolle. Denne prosessen skjer ikke på slump, men via høyst regulerte og presise kontrollsystemer, som det gjør i alle dyr. Tiden til rådighet for mutasjoner er heller knapp, for enklere organismer: noen få døgn. De enkleste organismer, (f.eks. en voksen sjøorm), har kun litt over 1000 celler. Likevel har den dGRNs av bemerkelsesverdig presisjon og kompleksitet. Utviklingen av kompleksiteten i embryoet kan måles i informasjonstermer, i følge Davidson.
Developmental gen regulatory networks (Dgrns) motstår mutasjonsendringer fordi de er hierarkisk organisert. Det innebærer at noen Dgrns kontrollerer andre Dgrns. I senteret av dette regulerende hierarkiet, er regulerende nettverk som spesifiserer akse og helhetlig kroppsplan under utviklingen. Selv små endringer i disse Dgrns vil medføre katastrofale virkninger på organismen. Likevel er det nettopp det som trengs om akse og global form på dyret skal variere: Nettopp de kretsløpene som ikke kan endres uten ødeleggelse som følge, må endre seg, om kroppsformen skal endres.
Konstruksjons begrensninger
Davidsons funn presenterer en dyptgående utfordring for tilstrekkeligheten til den neo-Darwinistiske mekanismen. Det å danne en ny type kropp krever ikke bare gener og proteiner, men nye dGRNs. Men en kan ikke lage nye dGRNs uten å endre eksisterende. Men det er nettopp dette som ikke er mulig, utenom multippelt koordinerte mutasjoner. I et hvert tilfelle har Davidsons arbeid vist oss at slike endringer, alltid har ført til katastrofale konsekvenser. Han gjør dette helt klart: 'I motsetning til klassisk evolusjonsteori, så kan ikke prosesser som drives fram av små endringer, bli tatt som modell for utviklingen av kropps-form for dyrearter'.. Dette kan ikke være overraskende, i følge Davidson, da neo-Darwinistisk syntese, stammer fra en premolekylær biologi, fokusert på populasjons genetikk og ..naturlig historie. Ingen av disse har noen inngang for dGRNs som styrer embryo-utvikling av kroppsformer..
Paleontologer forstår den Cambriske eksplosjonen som en plutselig geologisk tilsynekomst av nye livsformer. Å bygge disse kroppsuttrykkene krever nye utviklings program, både nye tidlig-handlende regulerende gener OG nye dGRNs. OM ingen av disse kan endres ved mutasjoner, uten å ødelegge eksisterende utviklingsprogram (og dermed dyreliv), så vil ikke naturlig utvalg ha noe å favorisere. Dermed vil utviklingen av nye dyreformer holde opp på det stadiet. Darwins tvil om fossilfunn i 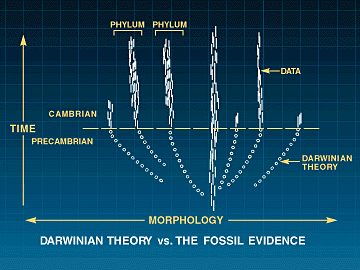 Cambrium er forsterket: Fossilene mangler, men i tillegg har vi et dyptgående problem med konstruksjons begrensinger. Problemet 'å bygge en ny form for dyreliv, ved gradvis å endre et tett integrert system av genetiske komponenter og deres produkter infiltrert i hverandre'. Ennå gjenstår mer formidable problemer for neo-Darwinismen:
Cambrium er forsterket: Fossilene mangler, men i tillegg har vi et dyptgående problem med konstruksjons begrensinger. Problemet 'å bygge en ny form for dyreliv, ved gradvis å endre et tett integrert system av genetiske komponenter og deres produkter infiltrert i hverandre'. Ennå gjenstår mer formidable problemer for neo-Darwinismen:
Epigenetisk/kontekst-avhengig informasjon
Fra Darwins Doubt kap. 14
Mange biologer tror ikke lenger at DNA styrer nærmest alt som skjer inne i cellen. Utviklingsbiologer spesielt, oppdager nå flere og flere måter som avgjørende informasjon til å bygge kroppsform blir tildelt via form og struktur av embryo-celler, inklusive informasjon fra både befruktet og ikke-befruktet egg.
Ordforklaringer: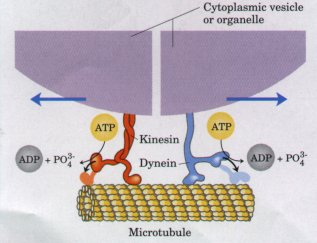
Centrioler: et par av svært små sylindriske organeller, nær kjernen i dyreceller. Involvert i utviklingen av spindelfibre i celledelingen.
Cytoskjelett: Internt i cellen: Cytoskjelettet virker som ankerfester og transportveier for makromolekyler og organeller, og deltar i organisering og romlig plassering av innholdet i cellene. Noen av oppgavene til cytoskjelettet er cytoplasmastrømning, flytting av kloroplaster avhengig (fra snl)
Eukaryote celler: Eukaryoter har en cellekjerne omgitt av en membran, samt bl.a. mitokondrier og mange andre organeller som mangler hos de prokaryote cellene (med kjerne). Bakterier er prokaryote, mens alle andre en- eller flercellede dyr er eukaryote.
Embryo-det tidlige foster
epi-genetisk- gresk forstavelse epi betyr over eller hinsides. Epigenetisk eller kontekstavhengig informasjon spiller avjørende rolle i prosessen som leder til et dyrs kropps-montering i fosterutviklingsperioden.
filament- tekstilfibre (filamentum-'tråd')
 Eukaryote celler har interne skjelett for å gi dem form og stabilitet. Disse skjelettene har ulike slag av fibre, inkludert mikrotuber . Strukturen og lokalisering av mikrotubene i cytoskjelettet influerer formen på og utviklingen av embryoet. Oppstillingen av mikrotuber innen embryo-celler, hjelper å distribuere essentielle proteiner som brukes under utvikling til spesifikke lokasjoner i cellen. Når de er dannet, utfører slike proteiner funksjoner kritiske for utviklingen av embryoet. Forutsetningen er at de blir levert til sine korrekte plasseringer, ved hjelp av preeksistente, presist formulerte oppstillinger av mikrotuber og cytoskjelett. Slik blir den presise arrangeringen av mikrotuber i cytoskjelettet en form for kritisk, strukturert informasjon.
Eukaryote celler har interne skjelett for å gi dem form og stabilitet. Disse skjelettene har ulike slag av fibre, inkludert mikrotuber . Strukturen og lokalisering av mikrotubene i cytoskjelettet influerer formen på og utviklingen av embryoet. Oppstillingen av mikrotuber innen embryo-celler, hjelper å distribuere essentielle proteiner som brukes under utvikling til spesifikke lokasjoner i cellen. Når de er dannet, utfører slike proteiner funksjoner kritiske for utviklingen av embryoet. Forutsetningen er at de blir levert til sine korrekte plasseringer, ved hjelp av preeksistente, presist formulerte oppstillinger av mikrotuber og cytoskjelett. Slik blir den presise arrangeringen av mikrotuber i cytoskjelettet en form for kritisk, strukturert informasjon.
Form: en distinkt utforming/fasong eller arrangement av kroppsdeler.
Mikrotuber: rørformede, 24 nm tykke fibrer, bygd opp av proteinene alfa- og beta-tubulin. Slike fibrer brukes ved celledelingen , ved intracellulær transport (se kinesin ) og i flimmerhår.
Morfogenetisk -genetikk som utvikler form
Morfologisk -formdannende organeller --er biologiske maskiner i nanoformat . Strukturer i celler som er omgitt av membran og har hver sine funksjoner.
stamceller - de allerførste celler i fosterutvikling, celler som i prinsippet kan bli til hva som helst i den påbegynte organisme.
Utviklingsbiologer befatter seg med dyrs utvikling fra befruktet eggcelle (zygogen) gjennom fosterutvikling og 'larve/unge-stadiet' til forplantningsdyktig individ.
Evolusjonsbiologer befatter seg med 'livsformenes evolusjon'
Utfordring for nåværende teoris fullstendighet
Nå har forskere funnet ut at dannelse av kroppsstruktur under fosterutvikling avhenger av et hierarkik av informasjonssystemer, på et langt høyere nivå enn DNA. Det samme hierarki må ha spilt en rolle i løpet av livets historie, når nye kroppsstrukturer er oppstått. Genetiske mutasjoner er langt fra noe riktig verktøy til den aktuelle jobben. Det kreves både genetisk og epigenetisk informasjon for å bygge en kroppsstruktur. Siden epigenetisk informasjon ikke er skjult i DNAet, kan ikke nye kroppsformer dannes ved DNA-mutasjoner.
Cellens form og struktur oppstår i hver ny generasjon, både som resultat av genprodukter og allerede eksisterende tredimensjonale strukturer, samt organisasjon som går i arv fra celler, cytoskjeletter og cellemembraner. Hverken proteiner eller de gener som koder for dem, kan hver for seg bestemme formen på enhetene de bygger. Logisk følge blir at naturlig seleksjon, heller ikke kan produsere de nye kroppsstrukturer som er oppstått i livets historie. Den handler bare ut fra genetisk variasjon og mutasjoner.
Strukturer der epigenetisk informasjon holder til, cytoskjelettets 'togbaner' og membran-mønstre, er mye mer omfangsrike enn cellekjernens DNA. Når en har forsøkt å endre slike strukturer, har det 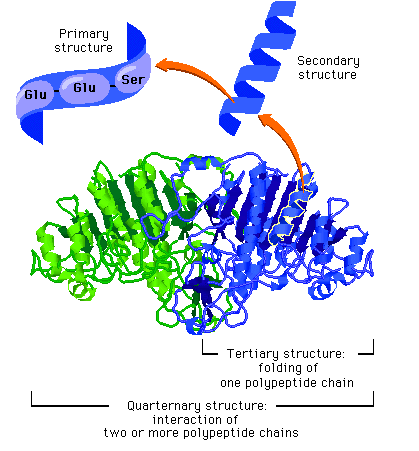 alltid ført til skadelige eller katastrofale konsekvenser. Eksperimenter har gjennomgående vist at ved omfattende endringer i epigenetisk informasjon, så har ikke embryoet en sjanse til å overleve i naturen, enn si å formere seg.
alltid ført til skadelige eller katastrofale konsekvenser. Eksperimenter har gjennomgående vist at ved omfattende endringer i epigenetisk informasjon, så har ikke embryoet en sjanse til å overleve i naturen, enn si å formere seg.
Påstand: Epigenetisk/kontekstuell informasjon spiller en avgjørende rolle i dannelse/sammensetning av dyr.
Eksempler som påviser at arv/endring av kroppsform avhenger av mer enn gener:

Saksområde: |
Påvisning: |
Begrunnelse: |
| Mønstre i cytoskjelettet |
Plassering avgjør- en form for kritisk informasjon |
Arves direkte |
| Mikrotubuler |
Lokasjon og arrangement avgjør arv |
Testbart |
| Ione-kanaler og elektromagnetiske felt |
Arrangering og plassering av ione-kanaler kan påvirke dyrets utvikling |
Kunstige felt kan forårsake celle-vandring. |
| Sukkermolekyler på cellemembran |
Plassering spiller kritisk rolle i funksjonen de har |
Testet ved snitt |
| Mål-punkter på celle-membran |
Spesifikk lokasjon og fordeling avgjør funksjon |
Plassering avgjøres ikke av genprodukt alene |
| Folding av proteiner |
Mange proteiner folder seg ulikt avhengig av omgivelsene |
Påvist i sammenlignende utviklingsbiologi |
| Morfogenetiske proteiner, regulerende gener og dGRNs |
Disse avhenger alle av plasseringen av spesifikke, informasjonsrike, tidligere cellestrukturer |
En har aldri opplevd at dGRNs er blitt endret med livskraftig avkom som følge |
| Cellestruktur |
Resultat av både genetikk og tidligere 3-dim-struktur og organisering i celle, cellemembran, og cytoskjelett |
Påvist ved forsøk at snitt i cellemembran arves direkte på encellede organismer |
Bilde -over: Ulike celletyper hos mennesket
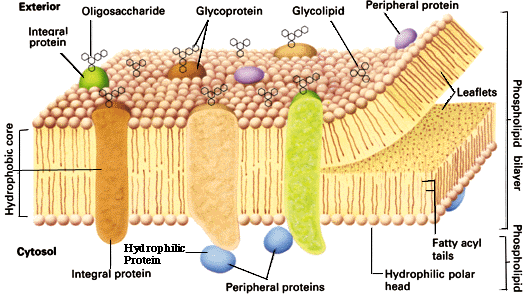 Cellemembranens målområder spiller en avgjørende rolle i utviklingen av embryoet, ved å tiltrekke molekyler til spesifikke plasser på indre celleoverflate. Når mange proteiner folder seg ulikt etter hvor de befinner seg i cellulær kontekst, så viser det kontekst-avhengig informasjon.
Cellemembranens målområder spiller en avgjørende rolle i utviklingen av embryoet, ved å tiltrekke molekyler til spesifikke plasser på indre celleoverflate. Når mange proteiner folder seg ulikt etter hvor de befinner seg i cellulær kontekst, så viser det kontekst-avhengig informasjon.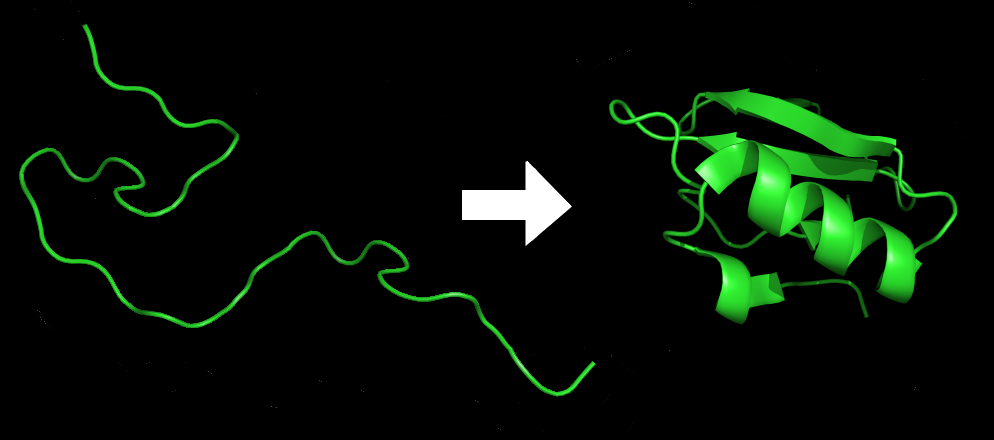
Genprodukter forsyner nødvendig, men ikke tilstrekkelig betingelser for utvikling av 3-dim struktur for celler, organer og kroppsplaner. Derfor blir neo-Darwinisme utilstrekkelig for å forklare dannelse av nye kropp-former. Darwinismen gir ‘survival of the fittest’, men ikke ‘origin of the fittest’. Darwin har ikke klart å gi forklaring på opphav til artene.
Epigenetiske mutasjoner?
Når flere blir oppmerksomme på de nye problemstillingene epigenetikken reiser, så spør en -av gammel vane, om ikke mutasjoner kan føre til større (makro) endringer her? Det viser seg at mutasjoner av epigenetisk informasjon, ikke er en realistisk vei for å genere nye livsformer:
1. Strukturene hvor epigenetisk informasjon er arvelige, membran-mønstre og cytoskjelettet, er mye større enn DNA-strenger. Av denne grunn er ikke strukturene så sårbare for endringer fra mange vanlige mutasjons-kilder på gener, slike som stråling eller kjemiske agenter.
 2. I den grad cellestrukturen er utsatt for endringer, er disse endringene i all hovedsak sannsynlige, å ha skadelige eller katastrofale konsekvenser. Sperman and Mangold utførte et eksperiment der de påtvang endringer i et viktig lager for epigenetisk informasjon, i løpet av embryo-perioden. Selv om resultatet hadde et interessant utseende, hadde det ikke sjanse til å leve opp, langt mindre formere seg.
2. I den grad cellestrukturen er utsatt for endringer, er disse endringene i all hovedsak sannsynlige, å ha skadelige eller katastrofale konsekvenser. Sperman and Mangold utførte et eksperiment der de påtvang endringer i et viktig lager for epigenetisk informasjon, i løpet av embryo-perioden. Selv om resultatet hadde et interessant utseende, hadde det ikke sjanse til å leve opp, langt mindre formere seg.
Den epigenetiske informasjonen som finnes i ulike celle-strukturer, er kritiske for utvikling av kroppsformer. F.eks. de regulerende proteinene laget av Hox-gener, og dGRNs avhenger alle plasseringen av spesifikk informasjonsrik og pre-eksisterende celle strukturer. Derfor vil endring av disse celle-strukturene etter all sannsynlighet ødelegge noe avgjørende viktig i tidlig embryo-stadium. Det er for mange enheter som er avhengig av epigenetisk informasjon, til at slike endringer kan ha positive, ja endog nøytrale effekter.
Darwins bok (Origin of Species) skulle f.o.f. være en forklaring på oppkomsten av nye arter . Han vedkjente at mønsteret for tilsynekomsten av dyreartene ikke skjedde i henhold til hans gradvise utviklings-bilde for livets historie. Mens han levde ble det ansett være et spørsmål om ufullstendighet i fossilmaterialet. Problemet har ikke kommet nærmere noen løsning, men nå er det et større og mer funamentalt problem som påvirker hele den darwinistiske konstruksjonen: Neo-darwinstisk metodologi kan verken gjøre rede for opprinnelsen til nødvendig genetisk eller epigenetisk informasjon for å produsere nye livsformer.
Kjente mikroevolusjonære prosesser kan gjøre rede for mindre endringer som sykliske variasjoner innen fink-bestanden på Galapagos, bakteriers resistanse mot antibiotika eller skifting av farge på nattsommerfugler. Men mange biologer argumenterer nå med at neo-Darwinistisk teori ikke gir tilstrekkelig grunn for opprinnelsen til nye kroppsformer, eller begivenheter som den Cambriske eksplosjonen.
F.eks. utviklings-biolog Keit S. Thompson har uttrykt tvil hvorvidt stor-skala morfologiske endringer, kan skyldes mindre endringer på gen-nivå. Biologene Gilbert, Opitz og Raff har forsøkte utvikle en ny evolusjonsteori som supplement til klassisk neo-Darwinisme, som de ikke finner kan gi tilstrekkelig forklaring på stor-skala makroevolusjonære endringer.
Det sier noe om omfanget av tvilen som er reist på dette området at så mange ledende biologer og palentologer stilt spørsmål ved tilstrekkeligheten til mekanismer ved neo-darwinismen og om opprinnelse til nye arter ut fra gen-mutasjoner spesielt: Gerry Webster og Brian Goodwin, Günter Theissen, Marc Kirschner, John Gerhart, Jeffey Schwartz, Douglas Erwin, Eric Davidson, Eugene Koonin, Simon Conway Morris, Robert Carroll, Gunter Wagner, Henz-Algert Beker og Wolf-Eckhart Lönnig, Stuart Newman og Gerd Müller, Stuart Kaufmann, Peter Stadler, Heinz Saedler, James Valentine, Giuseppe Sermonti, James Shapiro og Michael Lynch.. De som hevder at det bare er en liten håndfull fanatikere som utfordrer neo-darwinismen må nok utvide synsfeltet noe.
Referanser:
1. Schwarz, "Sudden Origin," 3
3. W. McGinnis og M. Kurziora, "The molecular Architects of Body Design," Scientific American 270 (1994): 58-66.
4. D. L. Lindsley et al, "Guide to Genetic Variations of Drosophila Melanogaster, Carnegie Institution of Washington, no. 627.
5. E. Davidson og D. Erwin, "An Integrated View of Precambrian Eumetazoan Evolution," Cold Spring Harbor Symposa on Quantitative Biology 74 (2009): 1-16.
7. E. Szathmary, "When the Means Do Not Justify the End," Nature 399 (1999): 745-746.
9. E. Szathmary, "When the Means Do Not Justify the End," Nature 399 (1999): 745-746.
Oversettelse samt utvalg av stoff og bilder ved Asbjørn E. Lund