Fossil Fredag: Ventetidsutfordringen for årsaksforholdet til darwinistiske forklaringer
Av Günter Bechly, 17. mai 2024. Oversatt herfra
 Denne fossile fredagen adresserer jeg den vanlige forespørselen om å gi en utvidet skriftlig versjon av forelesningene mine om ventetidsproblemet og artsparutfordringen (f.eks. På YouTube her), sammen med referanser til hovedstrøms vitenskapelige artikler som understøtter argumene mine. Jeg la også til et avsnitt om det kollapsende treproblemet som en ny utfordring for årsakstilstrekkeligheten til darwinistiske forklaringer på makroevolusjon. Siden ventetidsproblemet og .. artsparutfordringen .. begge er relatert til den veldig raske overgangen fra terrestriske grislignende forfedre som ligner på Indohyus eller Pakicetus til fullt marine delfinlignende hvaler som Dorudon og Basilosaurus, valgte jeg den ikoniske pakicetus attocki-fra tidlig Eocene fra Pakistan som dagens kjente fossil.
Denne fossile fredagen adresserer jeg den vanlige forespørselen om å gi en utvidet skriftlig versjon av forelesningene mine om ventetidsproblemet og artsparutfordringen (f.eks. På YouTube her), sammen med referanser til hovedstrøms vitenskapelige artikler som understøtter argumene mine. Jeg la også til et avsnitt om det kollapsende treproblemet som en ny utfordring for årsakstilstrekkeligheten til darwinistiske forklaringer på makroevolusjon. Siden ventetidsproblemet og .. artsparutfordringen .. begge er relatert til den veldig raske overgangen fra terrestriske grislignende forfedre som ligner på Indohyus eller Pakicetus til fullt marine delfinlignende hvaler som Dorudon og Basilosaurus, valgte jeg den ikoniske pakicetus attocki-fra tidlig Eocene fra Pakistan som dagens kjente fossil.
Som jeg har demonstrert i detalj i mange forelesninger og artikler (se Bechly 2024d), dokumenterer fossilprotokollen konsekvent en serie saltasjonelle overganger blitt kalt revolusjoner, eksplosjoner og 'Big Bang-er' av hovedstrøms evolusjonsbiologer, med god grunn (se også Bechly 2021, 2023a, Bechly & Meyer 2017). Dette fenomenet er allestedsnærværende i alle perioder av jordhistorien, i alle geografiske regioner, og i alle grupper av organismer, fra protister og planter til virvelløse dyr og virveldyr.
Disse diskontinuitetene i livets historie motsier ikke bare den darwinistiske kjerneprediksjonen av gradvis (dvs. en ansamling av små endringer over lengre tid), men reiser også et annet fatalt problem for muligheten for en hvilken som helst ikke-styrt prosess som tilstrekkelig forklaring på hovedoverganger i livets historie (makroevolusjon).
Bilde 1. Hvalens stamfar? (Pakicetus-attocki)
Ventetidsproblemet
Dette problemet kalles ventetidsproblemet og oppstår fra kombinasjonen av to felt i biovitenskapene som vanligvis anses å gi god støtte for darwinistisk evolusjon. Ett felt er paleontologi med de rike dataene fra en omfattende fossiljournal, som dokumenterer millioner av år med dyp tid og antatte overgangsformer (f.eks. Archeopteryx, Australopithecus og Tiktaalik) som er elegant forklart med vanlig avstamming. Dette etablerer utover rimelig tvil makroevolusjon. Det andre feltet er populasjonsgenetikk, som dokumenterer effekten av den neo-darwinistiske prosessen med naturlig (og seksuell) seleksjon som virker på tilfeldig genetisk variasjon, noe som fører til observerbare endringer i genfrekvensene.
 Et godt eksempel er opprinnelsen til medikamentresistens i populasjonen av bakterier i laboratorieeksperimenter. Dette etablerer ukontroversielt mikroevolusjon. Den generelle antagelsen er at en ekstrapolering av mikroevolusjon over lengre perioder tilstrekkelig forklarer makroevolusjon. Når vi virkelig kombinerer resultatene fra begge nevnte fagområder, kommer vi imidlertid til en alvorlig utfordring for den matematiske gjennomførbarheten av den neo-darwinistiske prosessen. Mens fossilprotokollen gir velbestemmede tidsvinduer som var geologisk tilgjengelige for større overganger for å utfolde seg, gir standard lærebokpopulasjonsgenetikk et teoretisk rammeverk og matematisk verktøysett for å faktisk å utføre matematikken. Når slike beregninger og/eller simuleringer kjøres med rimelige estimater av alle parametere, er det entydige resultatet alltid at de geologisk tilgjengelige tidsvinduene, som er etablert av fossilprotokollen for alle de viktigste overgangene i livets historie, er størrelses-ordener for korte til å imøtekomme ventetidene som kreves, for at de nødvendige genetiske endringene oppstår og spres i forfedre populasjonen. Derfor viser ventetidsproblemet med matematisk strenghet (Hössjer et al. 2021) at neo-darwinisme ikke er en levedyktig forklaring på makroevolusjon.
Et godt eksempel er opprinnelsen til medikamentresistens i populasjonen av bakterier i laboratorieeksperimenter. Dette etablerer ukontroversielt mikroevolusjon. Den generelle antagelsen er at en ekstrapolering av mikroevolusjon over lengre perioder tilstrekkelig forklarer makroevolusjon. Når vi virkelig kombinerer resultatene fra begge nevnte fagområder, kommer vi imidlertid til en alvorlig utfordring for den matematiske gjennomførbarheten av den neo-darwinistiske prosessen. Mens fossilprotokollen gir velbestemmede tidsvinduer som var geologisk tilgjengelige for større overganger for å utfolde seg, gir standard lærebokpopulasjonsgenetikk et teoretisk rammeverk og matematisk verktøysett for å faktisk å utføre matematikken. Når slike beregninger og/eller simuleringer kjøres med rimelige estimater av alle parametere, er det entydige resultatet alltid at de geologisk tilgjengelige tidsvinduene, som er etablert av fossilprotokollen for alle de viktigste overgangene i livets historie, er størrelses-ordener for korte til å imøtekomme ventetidene som kreves, for at de nødvendige genetiske endringene oppstår og spres i forfedre populasjonen. Derfor viser ventetidsproblemet med matematisk strenghet (Hössjer et al. 2021) at neo-darwinisme ikke er en levedyktig forklaring på makroevolusjon.
Ventetidene øker dramatisk hvis koordinerte mutasjoner i stedet for bare punkt- mutasjoner er nødvendige for en tilpasning. Koordinerte mutasjoner er definert som enhver kombinasjon av to eller flere sammenfallende genetiske endringer som er individuelt nøytrale, og bare i kombinasjon gir en fenotypisk adaptiv fordel som gjør at utvalg kan fungere. At slike koordinerte mutasjoner faktisk er nødvendige for mange tilpasninger, er empirisk kjent for å være tilfelle, og også tydelig fra det faktum at mange (til og med enkle) trekk viste seg å ha en polygen kontroll.
Bilde 2. Quinine-resistens
Så her er hvordan ventetidsproblemet er avledet:
*Evolusjonen er ment å fortsette ved tilfeldige mutasjoner og naturlig/seksuell seleksjon.
*Seleksjon kan bare fungere med mutasjoner med en positiv eller negativ adaptiv verdi.
*I det minste noen adaptive fordeler krever to eller flere koordinerte mutasjoner.
*Enhver evolusjonær tilpasning krever mutasjoner som må oppstå og spre seg i en popuasjon. Dermed har alle mutasjoner to tidsbegrensninger som hovedsakelig er avhengige av mutasjonshastighet, populasjonsstørrelse og generasjonstid: Ventetiden for at en mutasjon skal oppstå og ventetiden for fiksering av denne mutasjonen.
SPØRSMÅL: Gir livets historie tilstrekkelige ressurser for evolusjon til å imøtekomme disse ventetidene? Svaret kan vises å være negativt!
 Konklusjon: Neo-darwinisme tilbakevises matematisk som en levedyktig teori om makroevolusjon.
Konklusjon: Neo-darwinisme tilbakevises matematisk som en levedyktig teori om makroevolusjon.
Faktisk er ventetidsproblemet ikke bare et enkelt problem, men snarere et uløselig dilemma, fordi uansett hvordan du endrer den avgjørende parameteren for populasjonsstørrelse, vil det alltid øke en av de to ventetidene. Med store populasjonsstørrelser synker ventetiden for at en mutasjon skal oppstå, men fikseringstiden øker (det samme er tilfelle for nøytral evolusjon). Med små populasjonsstørrelser øker ventetiden for at en mutasjon skal oppstå, men fikseringstiden avtar. Dermed er det ingen enkel måte for evolusjon å jobbe rundt ventetidsproblemet, f.eks. ved hjelp av genetisk drift og grunnlegger (founder) effekten.
Den fantastiske kraften i argumentet fra ventetidsproblemet, er at det ikke er avhengige av uklare konsepter, men faktisk lar deg gjøre matematikken, basert på det godt stabiliserte matematiske apparatet til hovedstrøms populasjonsgenetikk. Alle nødvendige parametere er enten kjent empirisk, eller deres område kan rimelig estimeres ved sammenligning med nylige pårørende (f.eks. mutasjonshastigheter, effektive populasjonsstørrelser, generasjons omløps-tider, lengde på bindingssteder osv.). Fossilprotokollen og radiometrisk datering gir dataene for de tilgjengelige tidsvinduene.
Ventetidsproblemet for koordinerte mutasjoner ble først formulert av Bodmer (1970) og Karlin (1973) og senere mye omtalt i hovedstrøms populasjonsgenetisk litteratur (f.eks. Christiansen et al. 1998, Beerinkel et al. 2007, Schweinsberg 2008, Durrett et al. Det ble anerkjent som et argument for intelligent design av Behe og Snoke (2004, 2005) og Behe (2007, 2009), og senere utdypet i flere publikasjoner av ID -talsmenn (f.eks. Axe 2010, Meyer 2013, Lemaster 2018 ). Jeg er helt enig med disse intelligente design talsmennene, som vurderte dette problemet som å pålegge en betydelig grense for effektiviteten til tilfeldige søkeprosesser.
Bilde 3. Ventetidsproblematikken utfordrer gjennomførbarhet av nd-mekanismer
Michael Behe fremmet i sin bok The Edge of Evolution (2007) argumentet om at ventetiden for to koordinerte mutasjoner er uoverkommelig for den neo-darwinistiske evolusjonsmekanismen til å fungere. HEFEE brukte empiriske data om en faktisk ventetid for en koordinert mutasjon som formidlet klorokin-medisinresistens i malaria. Anvendelse av disse empiriske dataene om menneskelig evolusjon, ved bare å korrigere for mye lavpopulasjonsstørrelse og mye lengre generasjonstid, spådde en ventetid på 10^15 år, som er mange størrelsesordre lengter enn eksistensen av vårt univers! De hovedstrøms befolkningsgenetikerne Durrett & Schmidt (2008) kritiserte Behes argument med en matematisk modell og hevdet at hans beregnede ventetid på 10^15 år er urealistisk. Imidlertid resulterte deres egne beregninger også i en uoverkommelig ventetid på 216 millioner år for en enkelt koordinert mutasjon i menneskelig evolusjon, noe som stort overskrider det tilgjengelige tidsvinduet på bare 6 millioner år siden splittelsen av den menneskelige avstamningen og sjimpansen avstamning fra en felles stamfar. Når resultatene fra en teoretisk modellforskjell så sterkt avviker fra eksperimentelle data fra den virkelige verden, bør vi heller stole på data fra den virkelige verden, fordi hver modell nødvendigvis må gjøre visse forenklinger som kan innføre feil. Følgelig er Behes tall imnest nærmere de sanne grensene for den darwinistiske mekanismen enn de fra Durrett og Schmidt. Uansett, forbyr og tilbakeviser begge tallene gjennomførbarheten av en darwinistisk mekanisme for makroevolusjon.
Ved hjelp av en annen modell, anvendte Sanford et al. (2015) en datasimulering for å beregne ventetidene i menneskelig evolusjon basert på rimelige estimater for en forfedres hominin-populasjon på 10.000 individer og en generasjons omløpstid på 20 år. De kom til fikseringstid på 85 millioner år for en enkelt samavhengig mutasjon, og 1,5-15,9 millioner år for en enkelt spesifikk punktmutasjon. Dette er veldig oppsiktsvekkende og uoverkommelig, og overgår det faktum at den antatte 5% forskjellen i det menneskelige vs sjimpanse genom innebærer millioner av mutasjoner som måtte oppstå og bli fiksert innen 6 millioner år, siden antatt separasjon fra deres respektive linjer. Dette kommer på toppen av ventetids-problemet for koordinerte mutasjoner, men er et eget argument.
 Selv hovedstrøms-forskere kom til lignende resultater, som Charlesworth (2020), som fant ut at ventetiden for at en ny mutasjon skulle oppstå og øke i frekvens i en populasjon kan være avskrekkende lang, og Pfennig et al. (2010), som fant at "hvis produksjonen av nylig favoriserte fenotyper krever nye mutasjoner, kan ventetiden for slike mutasjoner være forhindrende lang og sannsynligheten for etterfølgende tap gjennom drift kan være høy" (se også Pfennig 2021)
Selv hovedstrøms-forskere kom til lignende resultater, som Charlesworth (2020), som fant ut at ventetiden for at en ny mutasjon skulle oppstå og øke i frekvens i en populasjon kan være avskrekkende lang, og Pfennig et al. (2010), som fant at "hvis produksjonen av nylig favoriserte fenotyper krever nye mutasjoner, kan ventetiden for slike mutasjoner være forhindrende lang og sannsynligheten for etterfølgende tap gjennom drift kan være høy" (se også Pfennig 2021)
For noen år siden ble et tverrfaglig forskningsprosjekt opprettet ved Biologisk institutt ved Research Branch of Discovery Institute, for å utforske ventetids-problemet og demonstrere at de nevnte eksemplene ikke er unntak, men representerer et generelt mønster. Dette samarbeidsprosjektet inkluderer Douglas Axe (molekylærbiolog), Günter Bechly (paleontolog og prosjektleder), Michael Behe (biokjemiker), Ann Gauger (molekylærbiolog) Ola Hössjer (matematiker), Paul Nelson (filosof av biologi), og Richard von Sternberg (evolusjonsbiolog).
Som første trinn og teoretisk bakkearbeid utviklet vi en sterkt forbedret ny modell, basert på de matematiske modellene til Durrett & Schmidt (2008) og Sanford et al. (2015) samt Behrens & Vingron (2010). Denne modellen bruker Markov-kjeder av Moran-typen (iterativ stokastisk minneløs prosess for endelige befolkning) og flere forskjellige løsninger og inkluderer også fenomener som tilbake-mutasjoner og stokastisk tunneling. Dette arbeidet ble publisert i to fagfellevurderte mainstream-vitenskapsutsalg (Hössjer et al. 2018, 2021).
Bilde 4. Mulig å være en intellektuelt tilfreds teist (O. Grasso)
I et andre trinn brukte vi denne nye modellen på eksemplet med hvalers opprinnelse (Bechly et al. under forbereels), som regnes som et "plakat-reklame for makroevolusjon" (TheWissen & Bajpai 2001). Tidligere hadde Richard von Sternberg allerede gjort grove papir-beregninger basert på modellen i Durrett & Schmidt (2008), og på veldig sjenerøsee estimater for en effektiv populasjon på 100 000 individer per generasjon og en generasjonstid på 5 år. Resultatet var en ventetid på 43,4 millioner år for et enkelt par koordinerte mutasjoner. Fossilregisteret viser at bare 4,5 millioner år var tilgjengelig mellom representanter for fortsatt vandrende stamgruppe (Himalayacetus, 53,5 Mya) og de første virkelig vannhvalene som Basilosauridae (49 MYA). Denne dyptgripende overgangen krevde kompleks 'rekonstruksjoner' som ...:
*Forlemmer forvandlet til finner, reduksjon av bakben og bekken, halen forvandlet til flyndre (inkl. virvlrt for vertikal bevegelse)
*Re-Orienteringav foster for sub-aquatisk fødsel (hale-først)
*Endring av brystkjertler for die under vann
*Omorganisering av nyrevev for inntak av saltvann
*Spesiell lungeoverflateaktive middel (Lungen må utvides raskt etter å ha kommet opp til overflaten)
*Intra-abdominal motstrøms varmeutvekslingssystem (testikler er inne i kroppen ved siden av musklene som genererer varme under svømming)
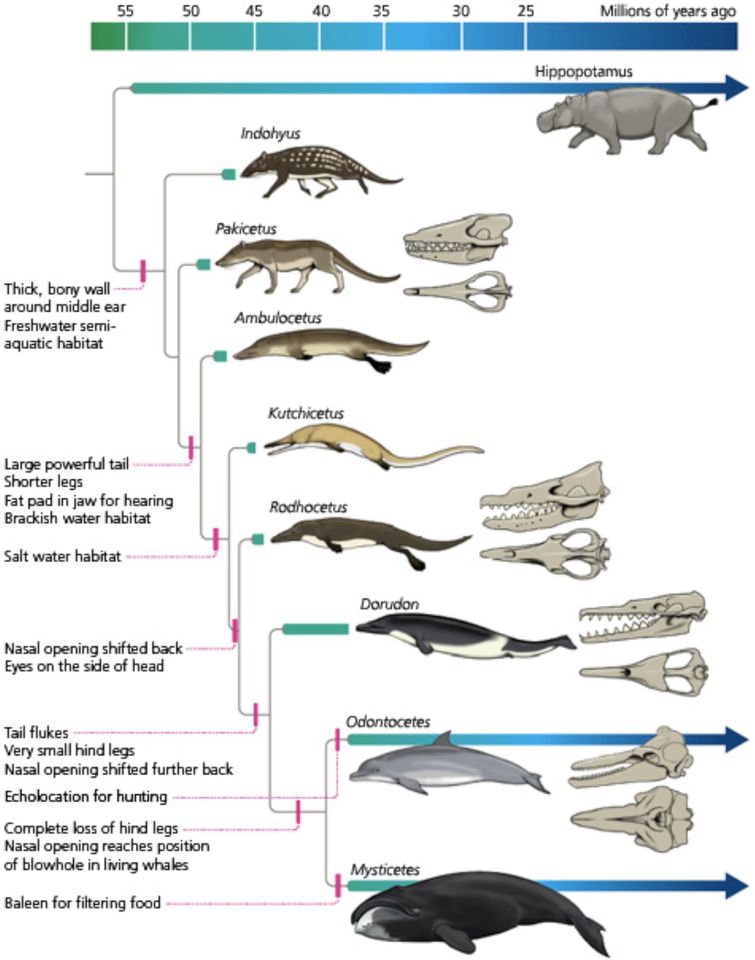 For vår nye studie fokuserte vi på overgangen til den amfibiske, men fortsatt firedoblet protoketider, som hadde en bakben-drevet modus for svømming, til fullt marine hvaler (Pelagiceti) med reduserte bakben og en hale-fluke-drivmote for svømming. Basert på en omfattende fossiljournal over de parafyletiske 'Walking Whales' (Protocidae, spesielt Georgiacetinae) og rester av de eldste basilosaurider fra Antarktis, fant vi at det tilgjengelige tidsvinduet for denne overgangen bare var 1,2 millioner år. Basert på moderne Evo-Devo-forskning demonstrerer vi også at koordinerte mutasjoner var nødvendige selv for tilsynelatende enkle trekk som reduksjon av bakben. Den matematiske modellen viser at den tilgjengelige tiden var enormt utilstrekkelig for å imøtekomme de nødvendige genetiske endringene ved en neo-darwinistisk mekanisme for naturlig seleksjon som virker på tilfeldige mutasjoner (Bechly et al. I prep.). Vi har allerede sendt dette manuskriptet til to hovedstrøms akademiske tidsskrifter, men de ble refusert i løpet av timer av redaktørene uten å bry seg med å sende det til vitenskapelige anmeldere. Tredje forsøk er på vei, men Darwinistisk tankepoliti gjør det veldig vanskelig å stille spørsmål ved hovedstrøms-konsensus, selv med harde data og god vitenskap. En tilknytning til forfatterne ved Discovery Institute eller et navn som er svartelistet som beryktet intelligent design -talsmann, er nok til å slippe løs Darwinismens vakthunder.
For vår nye studie fokuserte vi på overgangen til den amfibiske, men fortsatt firedoblet protoketider, som hadde en bakben-drevet modus for svømming, til fullt marine hvaler (Pelagiceti) med reduserte bakben og en hale-fluke-drivmote for svømming. Basert på en omfattende fossiljournal over de parafyletiske 'Walking Whales' (Protocidae, spesielt Georgiacetinae) og rester av de eldste basilosaurider fra Antarktis, fant vi at det tilgjengelige tidsvinduet for denne overgangen bare var 1,2 millioner år. Basert på moderne Evo-Devo-forskning demonstrerer vi også at koordinerte mutasjoner var nødvendige selv for tilsynelatende enkle trekk som reduksjon av bakben. Den matematiske modellen viser at den tilgjengelige tiden var enormt utilstrekkelig for å imøtekomme de nødvendige genetiske endringene ved en neo-darwinistisk mekanisme for naturlig seleksjon som virker på tilfeldige mutasjoner (Bechly et al. I prep.). Vi har allerede sendt dette manuskriptet til to hovedstrøms akademiske tidsskrifter, men de ble refusert i løpet av timer av redaktørene uten å bry seg med å sende det til vitenskapelige anmeldere. Tredje forsøk er på vei, men Darwinistisk tankepoliti gjør det veldig vanskelig å stille spørsmål ved hovedstrøms-konsensus, selv med harde data og god vitenskap. En tilknytning til forfatterne ved Discovery Institute eller et navn som er svartelistet som beryktet intelligent design -talsmann, er nok til å slippe løs Darwinismens vakthunder.
I 2009 var det en fremtredende offentlig debatt på Beverly Hills om intelligent designteori kontra darwinistisk evolusjonsteori, der Stephen Meyer og Richard Sternberg forsvarte ID -siden, mens den darwinistiske siden ble representert av skeptikeren Michael Shermer og den berømte palecetologen Dr. Donald Prothero, forfatter av boken Evolution: What the Fossil Say and Why it Matters. Da Sternberg presenterte ventetidsproblemet med eksemplet hval-opprinnelse, klarte ikke en ekspert som Protheroå virkelig forstå argumentet og viste seg å være uvitende for hvordan svare.
Bilde 5. Utviklingen måtte ha foregått i racer-fart
Et ekstra faktum som understreker ventetidsproblemet i tilfeller som hvalens opprinnelse (se Illustra Media 2015, Evolution News 2016, Long Story Short 2020) ble ironisk nok levert av Protheros egen forskning om den gjennomsnittlige levetiden (levetiden) av en større artiodactyl pattedyrart ( Prothero 2014), som vil omfatte hvaler og deres antatte forfedre fordi det antas at hvaler er nestet i og avledet fra artiodactyl-unge. Prothero fant at denne gjennomsnittlige artens livsperiode bare var rundt 4,1-4,39 millioner år. Fossilprotokollen innebærer derfor at overgangen forkynner en grislignende birebent hval-stamfar som ligner på Raoellidae (som Indohyus) og Pakicesidae, til en fullt marin delfinlignende Pelagicete-hval som Basilosauridae, skjedde innenfor levetiden til en enkelt art, eller til og med for Basilosauridae bare to millioner år, hvis det eldre estimatet på 51,5 millioner år for den eldste basilosauriden skal være riktig som vi antar (Bechly et al. I prep.). Dette utelukker selvfølgelig ikke muligheten for flere påfølgende spesifikasjonshendelser i løpet av denne tidsperioden, men representerer fortsatt et faktum som illustrerer den biologiske forkortelsen av denne store kroppsplanovergangen og anatomisk omvurdering innen bare 130 000-300 000 generasjoner.
Dette punktet gjelder også alle de andre brå opptredenene av biologisk nyhet som beskrevet i min andre artikkel , fordi ifølge standard læreboks visdom er den gjennomsnittlige levetiden til en dyreart 5-10 millioner år for marine virvelløse dyr (May et al. 1995: Tabell 1.1, Levinton 2001: Tabell 7.2), 3-10 millioner år for insekter (Hörnschemeyer et al. 2010, Penney & Jepson 2014: 198-199), og bare 2,3-4,43 millioner år for virveldyr som pattedyr (PRero 2014 ). Dermed tilsvarer et vindu på 5 millioner år omtrent en rekke på bare 1-2 forfedre etterkommer-arter med bare mindre forskjeller i til hverandre. Det er rett og slett for mye forandring på for kort tid til å bli rimelig forklart ved en ikke-styrt mekanisme!
Vi forventer derfor og spår at vi finner ventetidsproblemet å være et generelt problem for alle store kroppsplanoverganger i livets historie, med deres vanlige brå fremdukkinger (Bechly 2021, 2023a, Bechly & Meyer 2017), som for eksempel inkluderer: Opprinnelse til fotosyntesen; Opprinnelse til eukaryoter; Opprinnelsen til Ediacaran Biota (Avalon Eksplosion) og dyrefyla (kambrisk eksplosjon) som opprinnelsen til trilobitter fra ormlignende forfedre på mindre enn 13 millioner år (Daley et al. 2018, Bechly 2018); Opprinnelse til effektive øyne i leddyr, kefalopoder og virveldyr; Terrestrialisering av planter (embryofytter), leddyr (tracheae) og virveldyr (tetrapod lemmer); Opprinnelse til vinger hos insekter, pterosaurer, flaggermus og fugler (inkludert opprinnelsen til pennekurer fra filamentøse forløpere); Opprinnelse til sekundært marine virveldyr som ichthyosaurs, mosasaurer, manater og hvaler; Opprinnelse til ekkolokalisering i flaggermus og hvaler; Opprinnelse til komplekse nye reproduktive systemer (angiospermblomster, øyenstikker sekundære kopulerende apparater, insekt holometabolisme, amnioteegg, eutherisk morkake); Opprinnelsen til distinkte nye kroppsplaner (f.eks. slanger, skilpadder, flaggermus); og til og med opprinnelsen vår egen slekt Homo og av en kuleformet hjerne-kasse, korrelerte med den øvre paleolitiske 'kreative eksplosjonen' av symbolsk tenking i Homo sapiens.
Potensielle innvendinger til ventetidsproblemet
Ventetidsproblemet har vært målet for hånlig kritikk av anti-ID-talsmenn (f.eks. Moran 2016, Rasmussen 2021, Stern Cardinale 2022a, 2022b, Farina 2022, Hancock 2024), som hevdet at det er feilaktig og mislykkes i å utfordre darwinismen. Jeg adresserte en slik kritikk i flere artikler (Bechly 2022c, 2022d), og vi vil også gi en omhandlet teknisk respons i vårt kommende forsknings srtikkel (Bechly et al. under forberedelse). Hovedpoengene reist av kritikerne var:
 1.) Kritikere antydet ofte eksplisitt eller implisitt at ventetidsproblemet ikke er annet enn et pseudoproblem oppfunnet av Darwin-skeptikere og kreasjonister som ikke forstår hvordan evolusjonen virkelig fungerer. Dette er påviselig usant og unnlater å erkjenne at ventetidsproblemet har en lang historie og har blitt mye diskutert i hovedstrøms-vitenskap, spesielt populasjonsgenetikk (se over), inkludert utmerkede eksperter som Harvard evolusjonsbiolog Martin Nowak. Ventetidsproblemet spiller til og med en viktig rolle i kreftforskning (Beerinkel et al. 2007, Durrett et al. 2009).
1.) Kritikere antydet ofte eksplisitt eller implisitt at ventetidsproblemet ikke er annet enn et pseudoproblem oppfunnet av Darwin-skeptikere og kreasjonister som ikke forstår hvordan evolusjonen virkelig fungerer. Dette er påviselig usant og unnlater å erkjenne at ventetidsproblemet har en lang historie og har blitt mye diskutert i hovedstrøms-vitenskap, spesielt populasjonsgenetikk (se over), inkludert utmerkede eksperter som Harvard evolusjonsbiolog Martin Nowak. Ventetidsproblemet spiller til og med en viktig rolle i kreftforskning (Beerinkel et al. 2007, Durrett et al. 2009).
Bilde 6. Koordinerte mutasjoner
2.) De fleste kritikere anså den mektigste innvendingen for å være Texas skarpretter feilslutningen (f.eks. Moran 2016). De hevdet at naturen ikke går for spesifikke mutasjoner som et mål, men er helt tilfeldig og kunne ha forfulgt mange forskjellige måter å lykkes på. Kort sagt: Denne innvendingen hevder at du ikke kan anta et forhåndsbestemt mål. Dette argumentet mislykkes fordi det tvilsomt forutsetter eksistensen av mange mål, som er motsagt av sjeldenhetene av funksjon i søkeområdet for proteiner og av det vanlige fenomenet konvergens. Argumentet klarer heller ikke å erkjenne at livet ikke kan gi mulighet for perioder med mal-tilpasninger bare for å stige ned en lokal topp av det robuste egnethetslandskapet, for å utforske andre. I stedet må livet ytterligere tilpasse seg sin lokale egnethetstopp, som ber om spesifikke løsninger for spesifikke problemer. Ikke som noen gunstig mutasjon kan gjøre, eller som om treningslandskapet ville være flatt og homogent. En stemhval ville ikke ha nytte for en mutasjon som ville være gunstig for en stem-fugl, for eksempel å forbedre skjelettpneumatisiteten. Målene ble ikke vilkårlig pålagt, men blir gitt og begrenset av naturen. I datamodellene som ble brukt i publikasjonene våre om ventetidsproblemet, tillot vi også alternative mål og uklare mål, så ikke bare ett forhåndsbestemt bindingssted, som forhindrer en annen mulig kritikk. Generelt ser det ut til at denne innvendingen koker ned til den uvitenskapelige påstanden "Det kunne være x, slik at problemet forsvinner", som er en ikke-følge-påstand som ikke viser noe og ikke er testbar, fordi den ikke gjør noen spesifikke empiriske forutsigelser.
Forresten: I sammenheng med denne innvendingen antydet Stern Cardinal (2022a, 2022b) også en studie av Yona et al (2018) som viste at "tilfeldige sekvenser raskt utvikler seg til de novo -promotører". Dette er interessant, men refererer bare til en veldig enkel funksjon, som ble besvart av Miller (2022). Men enda viktigere denne studien er helt irrelevant for problemet med koordinerte mutasjoner, og er derfor bare en avledning. Den gjør ingenting for å få ventetidsproblemet til å forsvinne.
 3.) Noen kritikere klarte ikke å forstå begrepet koordinerte mutasjoner og kalte det til og med meningsløst. De antydet at hver enkelt mutasjon kan velges for. Dette ignorerer det enkle faktum at i koordinerte mutasjoner er hver individuell mutasjon nøytral og ikke kan selekteres for i prinsippet. Bare kombinasjonen av koordinerte mutasjoner har en seleksjonsverdi, som er hele poenget, og grunnen til at de ble kalt 'koordinerte mutasjoner' i utgangspunktet. Så ventetidsproblemet handler ikke om rene kombinasjonen av hvilken som helst form for mutasjoner. Denne unnlatelsen av å forstå konseptet med koordinerte mutasjoner fører ofte til veldig uvennlige feilrepresentasjoner av ventetidsproblemet, som angivelig hevder "det tar for lang tid å få nye ting", noe som selvfølgelig ikke er noe annet enn en karikatur- og stråmannsversjon av det virkelige problemet.
3.) Noen kritikere klarte ikke å forstå begrepet koordinerte mutasjoner og kalte det til og med meningsløst. De antydet at hver enkelt mutasjon kan velges for. Dette ignorerer det enkle faktum at i koordinerte mutasjoner er hver individuell mutasjon nøytral og ikke kan selekteres for i prinsippet. Bare kombinasjonen av koordinerte mutasjoner har en seleksjonsverdi, som er hele poenget, og grunnen til at de ble kalt 'koordinerte mutasjoner' i utgangspunktet. Så ventetidsproblemet handler ikke om rene kombinasjonen av hvilken som helst form for mutasjoner. Denne unnlatelsen av å forstå konseptet med koordinerte mutasjoner fører ofte til veldig uvennlige feilrepresentasjoner av ventetidsproblemet, som angivelig hevder "det tar for lang tid å få nye ting", noe som selvfølgelig ikke er noe annet enn en karikatur- og stråmannsversjon av det virkelige problemet.
Bilde 7. Formdannelse styres ikke at tilfeldige mutasjoner
4.) Noen kritikere hevdet at ventetidsproblemet innebærer at mutasjoner må forekomme i en spesifikk sekvens, og at det misforstår den parallelle naturen til darwinistiske prosesser. Dette er rett og slett usant og kanskje basert på en misforståelse av det tekniske uttrykket 'koordinert gen'. Faktum er at ingen ID -talsmann noen gang hevdet at ventetidsproblemet bare gjelder for en spesiell sekvens av mutasjoner. For ethvert sett med rimelige parametere vil ventetiden for koordinerte mutasjoner (dvs. mutasjoner som må forekommer sammen for å ha en seleksjonsverdi) være uoverkommelig, uavhengig av rekkefølgen på disse mutasjonene. Det som er sant er at ventetidsproblemet blir enda verre når nøytrale mutasjoner også må oppstå i en spesifikk sekvens. Imidlertid ville en spesifikk mutasjonssekvens i det virkelige liv bare være rimelig når hver enkelt mutasjon har en tilpasningsdyktig fordel og den ble selektert for, det ville være det motsatte av koordinerte mutasjoner. Derfor er dette argumentet en total avledning (red herring), ettersom det ikke gjelder koordinerte mutasjoner som det sentrale punktet i ventetidsproblemet.
5.) De fleste kritikere hevdet også at ventetidsproblemet ignorerer rekombinasjon, som ifølge Farina (2022) "grunnløst reduserer den dype evolusjonære fordelen" og "dramatisk akselererer akkumulering av fordeler." Stern Cardinale (2022b) var ettertrykkelig uenig med Hössjer et al. At "det er rimelig å tro at rekombinasjon kan ignoreres." Faktisk viser denne innvendingen heller en pinlig uvitenhet om den tekniske litteraturen om dette emnet, fordi påvirkningen av rekombinasjon på ventetidsproblemet har blitt studert av Christiansen et al. (1998), som har vist at: "Rekombinasjon senker ventetiden til en ny genotypisk kombinasjon først vises, men effekten er liten sammenlignet med mutasjonshastigheten og populasjonsstørrelsen." Selv om denne prosessen uten tvil tillater nøytrale mutasjoner (omtrent 75% av alle mutasjoner) å oppstå separat i en populasjon og å kombinere senere ved seksuell rekombinasjon, viste vi i våre fagfellevurderte artikler (Hössjer et al. 2018, 2021, Bechly et al). Det må også nevnes at selv om rekombinasjon kan fremskynde noen evolusjonære endringer mot et spesifikt mål, kan det også øke ventetiden ved å ødelegge forestående tilpasninger som er i ferd med å nå målet.
6.) Noen få kritikere hevdet at problemet bare er teoretisk, men ikke realistisk i biologiske termer, for eksempel fordi det ikke gjelder konkrete eksempler eller fordi koordinerte mutasjoner ikke er nødvendige. Vi vil ta opp den siste påstanden veldig grundig i vårt kommende papir, der vi bruker det teoretiske rammeverket på det konkrete eksemplet på hvalers opprinnelse. Vi vil også vise, basert på hovedstrøms Evo-Devo-data, at koordinerte mutasjoner faktisk er påkrevd. Dette antydes også av det faktum at til og med enkle trekk som hudfarge viste seg å være svært polygen, og dermed kontrollert av mange forskjellige gener og som sannsynligvis krever flere koordinerte alleliske substitusjoner (mutasjoner) av rette slag, for å ha en adaptiv effekt. Forresten: Ventetidsproblemet har blitt brukt på det konkrete eksemplet på menneskelig opprinnelse av Durrett & Schmidt (2008) og Sanford et al. (2015) med uoverkommelige resultater for Darwinistisk evolusjon (se over).
 7.) En annen mulig innvending kan være studien av Wilf & Ewens (2010), som antydet at det var god tid til evolusjonen. Denne studien har tydeligvis vært et svar på argumenter fra Darwin-kritikere, ettersom abstraktet begynner med setningen "Innvendinger til darwinistisk evolusjon er ofte basert på tiden som kreves for å utføre de nødvendige mutasjonene." Denne studien ble feiret av anti-ID-aktivister som en gjendrivning av et populært argument mot neo-Darwinistisk Evolusjon (Coyne 2010, Myers 2010). Imidlertid begikk denne studien den samme trivielle logiske feilen som Richard Dawkins gjorde med sitt berømte weasel-program (Dawkins 1986), ved introduksjon et teleologisk filter for riktige gjetninger for å komme frem til Hamlets sitat "Methinks it is like a weasel" (Witt 2016, Holloway 2022). Dessuten har studien ikke bare blitt adressert og tilbakevist i en fagfellevurdert artikkel av ID-talsmenn (Ewert et al. 2012, Luskin 2012), men dens matematiske konklusjoner ble også utfordret i en nyere studie ledet av evolusjonsbiologen Martin Nowak (Chatterjee et al. 2014), som bekreftet det grunnleggende punktet i ventetidsproblemet at "at tiden som kreves for å akkumulere flere mutasjoner i samme individuelle skalaer, med antallet, ikke logaritmisk men eksponentielt" (Miller 2018).
7.) En annen mulig innvending kan være studien av Wilf & Ewens (2010), som antydet at det var god tid til evolusjonen. Denne studien har tydeligvis vært et svar på argumenter fra Darwin-kritikere, ettersom abstraktet begynner med setningen "Innvendinger til darwinistisk evolusjon er ofte basert på tiden som kreves for å utføre de nødvendige mutasjonene." Denne studien ble feiret av anti-ID-aktivister som en gjendrivning av et populært argument mot neo-Darwinistisk Evolusjon (Coyne 2010, Myers 2010). Imidlertid begikk denne studien den samme trivielle logiske feilen som Richard Dawkins gjorde med sitt berømte weasel-program (Dawkins 1986), ved introduksjon et teleologisk filter for riktige gjetninger for å komme frem til Hamlets sitat "Methinks it is like a weasel" (Witt 2016, Holloway 2022). Dessuten har studien ikke bare blitt adressert og tilbakevist i en fagfellevurdert artikkel av ID-talsmenn (Ewert et al. 2012, Luskin 2012), men dens matematiske konklusjoner ble også utfordret i en nyere studie ledet av evolusjonsbiologen Martin Nowak (Chatterjee et al. 2014), som bekreftet det grunnleggende punktet i ventetidsproblemet at "at tiden som kreves for å akkumulere flere mutasjoner i samme individuelle skalaer, med antallet, ikke logaritmisk men eksponentielt" (Miller 2018).
Bilde 8. Dawkins kom med innrømmelser om Kambrium
8) En ganske forvirret innvending ble reist av den darwinistiske YouTuber Dapper Dinosaur (2022), som hevdet at boarders som mutasjonshastighet og generasjonstid ikke virkelig betyr noe, men at den virkelig avgjørende parameteren skulle være den fylogenetiske erstatningsraten (Argestad 2019). Problemet med denne innvendingen er at den fylogenetiske erstatningshastigheten ikke har noen betydning for ventetidsproblemet, noe som også fremgår av det faktum at ingen av de vanlige evolusjonsbiologene, som diskuterte ventetidsproblemet noen gang gidder å vurdere en slik parameter i deres tekniske publikasjoner. Et annet problem er at Heritages konsept om en 'fylogenetisk erstatningshastighet' aldri har blitt publisert riktig og derfor ikke er en eksisterende ting i vitenskapen (et Google -søk resulterer bare i en enkelt hit for et konferansesammendrag publisert på nettet). Kravet til fagfellevurdering ser noen ganger ut til å være en forslitt enveis gate i debatten mellom darwinisme og intelligent designteori.
9) Noen kritikere av ventetidsargumentet, som Stern Cardinale (2022b), angrep i stedet John Sanfords forskjellige argument om 30 millioner bokstavforskjeller mellom menneskelige og sjimpanse genomer til å være for mye til å bli fikset på bare 6 millioner år siden separasjonen av disse to linjene. Dette er selvfølgelig nok en avledning ettersom dette argumentet ikke har noe med ventetidsproblemet å gjøre. Når det er sagt, påpekte Stern Cardinale riktig at det reelle antallet skulle være omtrent halvparten av denne størrelsen, fordi forskjeller ville ha samlet seg i begge lineforløp, men han drev også grovt feilaktig fremstilling av Sanfords argument, idet han angivelig hevdet 30 millioner gunstige mutasjoner mens Sanford eksplisitt bare snakket om'bokstav forskjeller' (se f.eks. her). Siden Sanford et al. (2015) fant fikseringstider på 1,5-15,9 millioner år selv for punktmutasjoner, kan denne mengden sekvensforskjeller uten tvil være et gyldig problem, men den er forskjellig fra ventetidsproblemet for koordinerte mutasjoner som brukes av intelligente design talssmenn.
10.) Sist, men ikke minst, ble noen kritikere forundret over at forskningsartikler om ventetidsproblemet som er forfattet av ID-talsmenn på en eller annen måte måte kunne "snike seg inn i" fagfellevurderte tidsskrifter som den prestisjetunge tidsskriftet for teoretisk biologi. årsaken er så enkel som den skal være åpenbar: fordi det er god fagfellevurdert vitenskap og den vanlige sensuren av anti-ID-aktivister noenganger unnlater å sabotere publiseringen av ubehagelig forskning. Det høyden av hykleri når de samme menneskene, som presser redaktører til å avvise manuskripter eller utstede ansvarsfraskrivelse, deretter snur og hevder at ID-talsmenn ikke publiserer tingene sine i fagfellevurdert litteratur.
 Senest ble en mer interessant kritikk presentert av evolusjonsbiologen Zach Hancock (2024), som introduserte i en YouTube -video sin egen matematiske datasimulering for å løse og tilbakevise ventetidsproblemet. Generelt, ville dette være veien å gå, men kritikken hans lider fortsatt av flere fatale problemer:
Senest ble en mer interessant kritikk presentert av evolusjonsbiologen Zach Hancock (2024), som introduserte i en YouTube -video sin egen matematiske datasimulering for å løse og tilbakevise ventetidsproblemet. Generelt, ville dette være veien å gå, men kritikken hans lider fortsatt av flere fatale problemer:
Hancock rekapitulerer dessverre mange av de samme tilbakeviste argumentene, omtalt ovenfor.
Hancock ser ut til å være uvitende om at den matematiske modellen til Hössjer et al. (2021) gjorde det mulig å håndtere Texas Sharpshooter-innvendingen av et forhåndsbestemt mål. Faktisk tok vi opp Texas skarpskytter -innvendingen (uten å bruke dette navnet) i siste avsnitt i avsnitt 11.2 og i vedlegg D. Spesielt i vedlegg D (tabell 16) tok vi opp hvor lang tid det ville ta å endre uttrykket til m av M mulige gener.
Vi rapporterte ventetiden til de første m= 5 generene kommer til uttrykk i et pool på M \ i {5, 10, 15, 20, 25} mulige gener. For et egnethetslandskap, der tilbake- mutasjoner er tillatt, er ventetiden fremdeles astronomisk for M = 25 (den høyre kolonnen i tabell 16). Vi gjør den testbare spådommen om at ventetiden fremdeles vil være enorm for m = 5 og M = 100, 500, 1000, hvis egnethetsdalen er dyp nok, og befolkningsstørrelsen stor nok. Videre nevnte vi biologiske argumenter hvorfor M ikke kan være veldig stor (se den siste setningen i vedlegg D).
Hancocks simulering, som bruker SLiM3 -programvaren, adresserer bare tilfellet m = 1 gen. Dette ser ut til å være mest som svar på resultatene fra Sanford et al. (2015) studie. Det er imidlertid ikke overraskende i det hele at ventetiden for m = 1 -genet er ganske kort. Selv om disse nye simuleringene av Hancock er interessante, er de ikke i konflikt med resultatene fra Hössjer et al. (2021) og unnlater å tilbakevise ventetidsproblemet.
Bilde 9. Matematikk/statistikk kan benyttes til å motbevise hypoteser
Resten (utvidet evolusjonær syntese) finnes her.
Konklusjon
Jeg (Bechly) foreslår at ventetidsproblemet, .. , har nådd en terskel av motstridende bevis som krever en paradigmendring i biovitenskap og en gjennoplevning av teleologi. Uansett hvilket nytt paradigme som vil erstatte neo-darwinisme, må fordelen av en slik ny tilnærming estimeres ikke bare basert på deres empiriske støtte, årsakssammenheng, generell forklaringskraft, sannsynlighet og parsimonier, men også basert på deres evne til å løse nevnte problemer.
Dette innebærer på ingen måte noen fornektelse av godt stablet vitenskap som geologisk datering og felles avstamming med modifisering, men innebærer en nødvendighet for et nytt syn på mekanismene som fører til biologiske nyheter, som ikke viker unna muligheten for målrettede prosesser. I stedet for å la materistisk verdenssyn-skjevhet bestemme hvilke forklaringer som er tillatt og hvilke som blir betraktet som tabu, så skal det bare være de vitenskapelige empiriske dataene i stedet for dogmatiske verdensbilder, som bestemmer blant konkurrerende hypoteser i et søk etter beste forklaring, etter bevisene -uansett hvor det fører.
For Litteratur se slutten av originalartikkelen -lenke.
Oversettelse, via google-oversetter, og bilder ved Asbjørn E. Lund